Metilarea ADN-ului

Metilarea ADN-ului este un proces biologic prin care grupele metil sunt adiționate la molecula de ADN. Metilarea poate modifica activitatea unui segment de ADN fără a-i modifica secvența. Când este localizată într-un promotor al genei, metilarea ADN-ului acționează, de obicei, pentru a reprima transcripția genei respective. La mamifere, metilarea ADN-ului este esențială pentru dezvoltarea normală și este asociată cu o serie de procese cheie, precum: amprenta genomică, inactivarea cromozomului X, reprimarea elementelor transpozabile, îmbătrânirea și carcinogeneza.
Până în prezent, se cunosc două nucleobaze pe care are loc metilarea naturală, enzimatică, a ADN-ului: adenina și citozina. Bazele modificate sunt N6-metiladenina,[1] 5-metilcitozina și N4-metilcitozina.[2]
| Baza nemodificata |  | 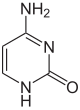 | ||||||
| Adenina, A | Citozina, C | |||||||
| Formele metilate |  | 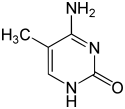 | 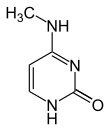 | |||||
| N6-metiladenină, 6mA | 5-metilcitozină, 5mC | N4-metilcitozină, 4mC | ||||||
| N.B. În cazul citozinei, numerotarea se realizează în sensul contrar acelor de ceasornic, începând de la atomul de azot poziționat la ora 6. În cazul adeninei, numerotarea se în același mod, însă începând de la atomul de azot poziționat la ora 2. Pentru conveniență, în locul denumirilor complete, se pot folosi prescurtările: 6mA, 5mC și 4mC. | ||||||||
Metilarea citozinei este larg răspândită atât la eucariote, cât și la procariote, chiar dacă rata de metilare variază semnificativ între specii: 14% din citozine sunt metilate la Arabidopsis thaliana, între 4% și 8% la Physarum,[3] 7,6% la șoarecele de casă, 2,3% la Escherichia coli, 0,03% la Drosophila; metilarea este nedetectabilă în Dictyostelium;[4][5] și practic absentă (metilare între 0,0002% și 0,0003%) la genul Caenorhabditis[6] sau la ciuperci precum Saccharomyces cerevisiae și S. pombe (dar nu și la N. crassa).[7][8] :3699 Metilarea adeninei a fost observată în bacterii, plante și, recent, în mamifere,[9][10] dar a fost mai puțin cercetată.
Metilarea citozinei pentru a forma 5-metilcitozină are loc în aceeași poziție (5) pe inelul pirimidinic unde este localizată gruparea metil a timinei; aceeași poziție distinge timina de baza analogă a ARN-ului uracilul, care nu are gruparea metil. Dezaminarea spontană a 5-metilcitozinei duce la formarea de timină. Aceasta are ca rezultat o nepotrivire de baze azotate T:G (timina mereu se împerechează cu adenina, și guanina mereu se împerechează cu citozina). Mecanismele de reparare ale ADN-ului corectează nepotrivirea înapoi la perechea originală C:G; alternativ, ele pot înlocui G cu A, transformând perechea T:G într-o pereche T:A, schimbând efectiv o bază și introducând o mutație. Această bază greșită încorporată nu va fi corectată în timpul replicării ADN-ului. Dacă celula intră în ciclul celular, mutația devine permanentă. Utilizarea aproape universală a timinei exclusiv în ADN, și a uracilului exclusiv în ARN, poate să fi evoluat ca un mecanism de control al erorilor, pentru a facilita îndepărtarea bazelor azotate de uracil generate de dezaminarea spontană a citozinelor.[11] S-a considerat că metilarea ADN-ului, precum și multe dintre metiltransferazele contemporane, au evoluat din activitatea primitivă de metilare a ARN-ului, ipoteza fiind susținută de o serie de dovezi.[12]
La plante și alte organisme, metilarea ADN-ului se găsește în trei contexte de secvențe: CG (sau CpG, unde p semnifică legătura fosfodiesterică între C și G), CHG sau CHH (unde H poate fi: A, T sau C). La mamifere însă, metilarea ADN-ului se găsește aproape exclusiv în dinucleotidele CpG, citozinele de pe ambele catene fiind de obicei metilate. Metilarea non-CpG poate fi totuși observată în celulele stem embrionare,[13][14][15] având rol în dezvoltarea neuronală.[16] În plus, metilarea non-CpG a fost observată și în celulele progenitoare hematopoietice, în special în secvența CpApC.[17]
Funcția conservată de metilare a ADN-ului

Profilul de metilare al ADN-ului vertebratelor este diferit de cel al altor organisme. La mamifere, aproximativ 75% din dinucleotidele CpG sunt metilate în celulele somatice,[18] metilarea ADN-ului aparând ca o stare implicită.[15][19] În contrast, genomul majorității plantelor, nevertebratelor, ciupercilor sau protistelor prezintă modele de metilare „mozaicate”, fiind caracterizat prin alternanța domeniilor metilate și nemetilate.[20][21]

Rata ridicată de metilare a secvențelor CpG din genomul mamiferelor are un cost evolutiv, deoarece crește probabilitatea mutațiilor spontane. Pierderea grupărilor amino are loc cu o frecvență ridicată pentru citozine, având consecințe diferite în funcție de metilarea acestora. Astfel, citozinele metilate, prin dezaminare spontană, formează timine; prin urmare, dinucleotidele CpG se transformă în mod constant la dinucleotide TpG, ceea ce este evidențiat de subreprezentarea dinucleotidelor CpG în genomul uman (acestea apar la doar 21% din frecvența așteptată).[22] Dezaminarea citozinelor nemetilate însă, transformă acestea în uracili care sunt rapid recunoscuți, procedându-se la repararea ADN-ului de către celulă.
Insule CpG
La mamifere, singura excepție pentru această epuizare globală a secvențelor CpG rezidă într-o categorie specifică de secvențe bogate în GC, și CpG, numite insule CpG, care rămân, în general, nemetilate și, prin urmare, păstrează conținutul originar de CpG.[23] Insulele CpG sunt definite pe baza următoarelor criterii:
1) au o lungime mai mare de 200 bp,
2) un conținut de secvențe G+C în proporție mai mare de 50% din total,
3) raportul dintre CpG observat și așteptat mai mare de 0,6, deși uneori sunt folosite și alte definiții.[24]
Excluzând insulele ce se repetă, există aproximativ 25.000 de insule CpG în genomul uman, dintre care 75% au o lungime mai mică de 850 bp.[22] Ele sunt unități de regulare majore și aproximativ 50% din insulele CpG sunt situate în regiunile promotoare ale genelor, în timp ce alte 25% se află în corpurile genice, servind adesea ca promotori alternativi. Reciproc, aproximativ 60-70% dintre genele umane au o insulă CpG în regiunile lor promotoare.[25][26] Majoritatea insulelor CpG sunt nemetilate și facilitează interacțiunea cu histonele subiacente, cum ar fi metilarea H3K4. În țesuturile somatice, doar 10% din insulele CpG sunt metilate, majoritatea fiind situate în regiuni intergenice și intragenice. [ <span title="This claim needs references to reliable sources. (March 2024)">necesită citare</span> ]
Reprimarea promotorilor denși CpG
Metilarea ADN-ului a fost, probabil, prezentă într-o oarecare măsură la eucariotele timpurii. Practic, în fiecare organism analizat, metilarea la nivelul regiunilor promotoare se corelează negativ cu expresia genelor respective.[20][27] Promotorii CpG-densi ai genelor transcrise activ nu sunt niciodată metilați, dar, pe de altă parte, există și genele silențioase din punct de vedere transcripțional care nu poartă promotori metilați. La șoarece și la om, aproximativ 60-70% dintre gene au o insulă CpG în regiunile lor promotoare și majoritatea acestor insule CpG rămân nemetilate, independent de activitatea transcripțională a genei, atât în celulele diferențiate, cât și în cele nediferențiate.[28][29] În timp ce metilarea insulelor CpG este neîndoielnic legată de reprimarea transcripțională, metilarea promotorilor săraci în CG rămâne neclară; ar putea fi irelevantă din punct de vedere funcțional.[30]
Metilarea ADN-ului poate afecta transcrierea genelor în două moduri. În primul mod, metilarea în sine poate împiedica fizic legarea proteinelor transcripționale la genă,[31] și în al doilea mod, și probabil mai important decât primul, ADN-ul metilat se poate lega de proteine ce conțin domeniul de legare a metil-CpG (MBD). Proteinele MBD recrutează apoi proteine suplimentare în locus-ul respectiv, cum ar fi histon-deacetilazele și alte proteine de remodelare a cromatinei cu efect asupra histonelor, formând astfel o cromatină compactă, inactivă, numită <i>heterocromatină</i>. Această legătură între metilarea ADN-ului și structura cromatinei este foarte importantă. De exemplu, anumite protooncogene sunt în acest fel oprite de la transcripție (i.e. devin gene silențioase). A se vedea sindromul Rett.
Reprimarea elementelor transpozabile
Metilarea ADN-ului este un mijloc important de represiune transcripțională, cel puțin în contextul promotorilor denși CpG. Reprimarea transcripțională a genelor pare limitată la clase foarte specifice de gene, care trebuie să rămână permanent silențioase, în aproape toate țesuturile. Deși metilarea ADN-ului nu deține flexibilitatea necesară pentru reglarea fină a transcripției genelor, stabilitatea oferită este ideală pentru asigurarea reprimării transcripționale permanente a elementelor transpozabile.[32] Controlul transpozonilor reprezintă una dintre cele mai vechi funcții ale metilării ADN-ului, regăsindu-se la animale, plante și mai mulți protiști.[33] Unii sugerează că metilarea ADN-ului a evoluat tocmai pentru acest scop.[34]
Expansiunea genomului
Se cunoaște că metilarea ADN-ului elementelor transpozabile a fost un mijloc de expansiune a genomului. Cu toate acestea, motorul evolutiv pentru expansiunea genomului rămâne necunoscut. Există o corelație clară între dimensiunea genomului și prezența CpG, ceea ce sugerează că metilarea ADN-ului elementelor transpozabile a dus la o creștere vizibilă a masei ADN-ului.[35]
Metilarea corpurilor genelor frecvent transcrise
O funcție care pare chiar mai bine conservată decât reprimarea transpozonilor, este reprezentată de corelarea pozitiă cu expresia genică. La aproape toate speciile unde este prezentă metilarea ADN-ului, aceasta se găsește în special în corpul genelor frecvent transcrise.[20][27] Funcția aceasta nu este bine înțeleasă. O serie de dovezi sugerează că ar putea regla matisarea[36] și suprima activitatea unităților transcripționale intragene (promotori criptici sau elemente transpozabile).[37] Metilarea corpului genei pare strâns legată de metilarea H3K36. La drojdii și mamifere, metilarea H3K36 este abudentă în corpurile genelor frecvent transcrise. În drojdie, H3K36me3 recrutează enzime, precum histon-deacetilaze, pentru a comprima cromatina, prevenind activarea siturilor de pornire criptice (v. mai sus Reprimarea promotorilor denși CpG).[38] La mamifere, enzimele DNMT3a și DNMT3b se leagă de H3K35me3 prin intermediul domeniului PWWP. [ <span title="This claim needs references to reliable sources. (March 2024)">necesită citare</span> ]
La mamifere

În timpul dezvoltării embrionare
Există o serie de modele de metilare a ADN-ului generațional-recurente în cursul dezvoltării unui mamifer. Aproape toate metilările de la părinți sunt șterse, mai întâi în timpul gametogenezei și, din nou, în embriogeneza timpurie, demetilarea și remetilarea având loc de fiecare dată. Demetilarea din embriogeneza timpurie are loc în perioada preimplantare în două etape – inițial în zigot, apoi în timpul primelor câteva cicluri de replicare embrionară a morulei și blastulei. Primul val de metilare are loc apoi în timpul etapei de implantare a embrionului, insulele CpG fiind însă protejate. Acest lucru are ca rezultat o represiune globală și permite ca genele de mentenanță să fie exprimate în toate celulele. În etapa post-implantare, modelele de metilare sunt specifice fiecărui țesut și stadiu aferent de dezvoltare, modificările definind fiecare tip celular, și menținându-se pe termen lung. Studiile asupra dezvoltării membrelor de șobolan din timpul embriogenezei au ilustrat natura dinamică a metilării ADN-ului. În acest context, au fost observate variații în metilarea globală a ADN-ului, în funcție de stadiul de dezvoltare și factorii de mediu, evidențiind reglarea complexă a proceselor de metilare din timpul organogenezei și implicațiile potențiale pentru strategiile de medicină regenerativă.[39]
Deși metilarea ADN-ului în sine nu este necesară pentru represiunea transcripțională, funcția sa este cu siguranță de blocare a transcripției prin schimbări conformaționale. În special, metilarea ADN-ului este esențială pentru menținerea represiunii transcripționale mono-alelice în contextul imprimării genomice și al inactivării cromozomului X.[40][41] În aceste cazuri, alelele silențioase diferă de cele exprimate prin statusul metilării, iar pierderea/dereglarea acestor statuturi are ca rezultat pierderea caracterelor imprimării și expresia alelelor normal inactive de pe cromozomii X. Metilarea ADN-ului pare absolut necesară în celulele diferențiate, deoarece eliminarea oricăreia dintre cele trei ADN metiltransferaze competente are ca rezultat moartea intrauterină sau post-partum. În schimb, metilarea ADN-ului este dispensabilă în tipurile de celule nediferențiate, cum ar fi masa celulară internă a blastocistului, celulele germinale primordiale sau celulele stem embrionare.
Datorită fenomenului de imprimare genomică, genomii matern și patern sunt marcați diferit, trebuind reprogramați corespunzător de fiecare dată când trec prin linia germinativă. Prin urmare, în timpul gametogenezei, celulele germinale primordiale trebuie să aibă modelele originare (biparentale) de metilare a ADN-ului șterse, remetilarea făcându-se în funcție de sexul persoanei transmițătoare. După fertilizare, genomurile patern și matern sunt din nou demetilate și remetilate (cu excepția regiunilor metilate diferențial asociate cu genele imprimate). Această reprogramare este probabil necesară pentru totipotența embrionului nou format și ștergerea modificărilor epigenetice dobândite.[42]
În cancer
În multe procese fiziopatologice, cum ar fi cancerul, insulele promotoare CpG ajung hipermetilate, ceea ce determină represiunea transcripțională a genelor respective care poate fi moștenită de celulele fiice.[43] Modificările în metilarea ADN-ului au fost recunoscute ca fiind componente importante ale dezvoltării cancerului. Hipometilarea, în general, apare mai devreme și este legată de instabilitatea cromozomială și pierderea imprimării genomice, în timp ce hipermetilarea este asociată cu promotorii și poate apărea (și) secundar reprimării genelor supresoare de tumori (i.e. gene care au rol în supresia tumorală), fiind o țintă pentru terapia epigenetică.[44] În contextul importanței metilării ADN-ului în procesele de dezvoltare și diferențiere ale organismului, datorită dereglării acestui proces pot apărea patologii de dezvoltare. În mugurii membrelor de șobolan, diferitele stagii de condrogeneză sunt asociate cu diferite profiluri de metilare.[39]
Hipometilarea globală a fost, de asemenea, dovedită a fi implicată în dezvoltarea și progresia cancerului prin diferite mecanisme.[45] De obicei, există hipermetilarea genelor supresoare de tumori și hipometilarea oncogenelor.[46]
Expresia alterată a microARN-urilor are și ea efect de activare/represiune a multor gene responsabile în progresia către cancer (vezi rolul microARN-urilor în cancer). Însă această alterare la rândul său are loc prin hiper-/hipometilarea situsurilor CpG din insulele CpG în promotorii (i.e. promotorii denși CpG) care controlează transcripția microARN-urilor.
Reprimarea genelor de reparare a ADN-ului prin metilarea promotorilor denși CpG aferenți pare a fi deosebit de importantă în progresia către cancer (vezi metilarea ADN-ului în cancer).
În ateroscleroză
Polimorfismele de metilare ale ADN-ului pot fi utilizate ca biomarkeri precoce ai aterosclerozei, deoarece sunt prezente înainte de apariția leziunilor, putând constitui un instrument pentru detectarea și prevenirea riscurilor.[47]
Două tipuri de celule afectate de polimorfismele de metilare ale ADN-ului sunt monocitele și limfocitele, care devin hipometilate. Mecanismul probabil pentru această hipometilare este determinat de nivelul crescut de homocisteină care cauzează hiperhomocisteinemie, un factor de risc pentru bolile cardiovasculare. Nivelurile plasmatice ridicate de homocisteină inhibă ADN-metiltransferazele, ceea ce provoacă hipometilarea. Hipometilarea ADN-ului afectează gene care modifică proliferarea celulelor musculare netede, provoacă disfuncția celulelor endoteliale și cresc mediatorii inflamației, toate aceste aspecte fiind esențiale în formarea leziunilor aterosclerotice.[48] Pe lângă acestea, nivelurile crescute de homocisteină pot avea ca rezultat și hipermetilarea promotorilor CpG denși ai genei receptorului de estrogen alfa (ERα), determinând infrareglarea acestuia.[49] ERα protejează împotriva aterosclerozei datorită acțiunii sale de supresor a proliferării celulelor musculare netede.[50] Hipermetilarea promotorului ERα permite astfel celulelor musculare netede intimale să prolifereze excesiv și să contribuie la dezvoltarea leziunii aterosclerotice.[51]
O altă genă care suferă o schimbare a statusului de metilare în ateroscleroză este transportorul monocarboxilat (MCT3), care produce o proteină responsabilă de transportul lactatului și a altor corpi cetonici din multe tipuri de celule, inclusiv din celulele musculare netede de la nivel vascular. La pacienții cu ateroscleroză, există o creștere a metilării insulelor CpG din exonul 2, ceea ce scade expresia proteinei MCT3. Infrareglarea MCT3 afectează transportul lactatului și crește semnificativ proliferarea celulelor musculare netede, ceea ce contribuie și mai mult la leziunea aterosclerotică. Un experiment ex vivo a arătat că utilizarea agentul de demetilare Decitabină (5-aza-2-deoxicitidină) induce expresia MCT3 într-o manieră dependentă de doză, deoarece insulele CpG hipermetilate din exonul 2 au devenit demetilate după tratament. Astfel, Decitabina ar putea servi ca un nou agent terapeutic pentru tratarea aterosclerozei, deși până acum nu au fost efectuate studii pe oameni.[52]
În insuficiența cardiacă
Există modificări epigenetice specifice și în insuficiența cardiacă. Acestea variază în funcție de etiologia bolii. De exemplu, în insuficiența cardiacă ischemică, modificările de metilare ale ADN-ului sunt asociate cu expresia genelor ce au rol în schimbarea metabolismului inimii ca urmare a ischemiei.[53] Alte forme ale insuficienței cardiace (de exemplu, cardiomiopatie diabetică) și comorbidități (de exemplu, obezitatea) trebuie explorate pentru a vedea cât de comune sunt aceste mecanisme. S-a determinat că aceste modificări de metilare sunt influențate și de factori rasiali și socioeconomici.[54]
În îmbătrânire
La oameni și la alte mamifere, nivelurile de metilare a ADN-ului pot fi utilizate pentru a estima cu exactitate vârsta țesuturilor și a celulelor, funcționând ca un ceas epigenetic precis.
Un studiu longitudinal al copiilor gemeni a arătat că, între 5 și 10 ani, divergențele modelelor de metilare s-au datorat influențelor de mediu, nu celor genetice.[55] Există o pierdere globală a metilării ADN-ului odată cu îmbătrânirea.[46]
Într-un studiu care a analizat metilomii (i.e. metilarea tuturor citozinelor dintr-o oarecare secvență de ADN) ADN-ului celulelor T CD4 + la un nou-născut, un individ de 26 de ani și un individ de 103 ani, s-a observat că pierderea metilării este proporțională cu vârsta.[56] Insulele CpG hipometilate observate în ADN-urile centenare, în comparație cu nou-născuții, acoperă toate compartimentele genomice (promotori, regiuni intergenice, introni și exoni).[56] Cu toate acestea, unele gene devin hipermetilate odată cu vârsta, inclusiv genele pentru receptorul de estrogen, p16 și IGF-2.[46]
În exercițiu
S-a demonstrat că exercițiile de mare intensitate reduc gradul de metilare al ADN-ului în mușchii scheletici.[57] Metilarea promotorului PGC-1α și PDK4 a fost redusă imediat după exerciții de intensitate mare, în timp ce metilarea PPAR-γ nu a fost redusă decât la trei ore după exercițiu.[57] În același timp, șase luni de exerciții fizice la bărbații de vârstă mijlocie anterior sedentari au dus la o metilare crescută la nivelul țesutului adipos.[58]
În diferențierea celulelor B
Un studiu care a investigat metilomul celulelor B de-a lungul ciclului lor de diferențiere, a arătat că există o hipometilare în toate stadiile. Cea mai mare diferență de metilare este între etapele celulelor B ale centrului germinal și ale celulelor B cu memorie. În plus, acest studiu a arătat că există o similitudine în profilul de metilare al ADN-ului între tumorile celulelor B (limfoame) și celulele B cu viață lungă.[17]
În creier
Două recenzii de literatură de specialitate au constatat că modificările de metilare ale ADN-ului din neuronii creierului sunt importante pentru învățare și memorare.[59][60] Procesul de învățare prin asociere cu răspunsul la frică (modelul Pavlovian) la animale, cum ar fi șoarecii și șobolanii, este rapid și este extrem de robust în crearea amintirilor.[61] La șoareci[62] și la șobolani[63] condiționarea fricii contextuale, în decurs de 1-24 de ore, este asociată cu metilările câtorva mii de citozine din ADN-ul genelor neuronilor din hipocamp. La douăzeci și patru de ore după un astfel de eveniment, 9,2% dintre genele din neuronii hipocampului de șobolan sunt metilate diferențial.[63] La șoarecii examinați la patru săptămâni după condiționare, metilările și demetilările de la nivelul hipocampului au revenit la profilurile inițiale, în schimb metilări și demetilări diferențiale substanțiale ale insulelor CpG au avut loc în neuronii corticali în timpul procesului de mentenanță al memoriei, și au existat 1223 de gene metilate diferențial în cortexul cingulat anterior.[62] Aceasta demonstrează faptul că hipocampul este necesar pentru formarea amintirilor, dar amintirile nu sunt stocate acolo. Mecanismele care ghidează metilările și demetilările noi din hipocamp din timpul memorizării au fost rezumate într-un studiu din 2022.[64] Aceste mecanisme de metilare/demetilare dau naștere la noi modele de expresia a ARN-ului mesager. Aceste noi ARN-uri mesager au fost apoi transportate la nivelul sinapselor neuronale, unde pot fi translatate în proteine.[64] Modificările active în metilarea și demetilarea ADN-ului neuronal par să acționeze în calitate de controlori ai scalării sinaptice și ai traficului de receptori de glutamat cu rol în procesele de învățare și memorare.[59]
ADN-metiltransferazele (la mamifere)

În celulele mamiferelor, metilarea ADN-ului are loc în principal la poziția C5 a dinucleotidelor CpG, fiind realizată prin două procese generale enzimatice - metilarea de întreținere și metilarea de novo.[65]
Activitatea de întreținere a metilării păstrează metilarea ADN-ului după fiecare ciclu celular de replicare. Fără ADN-metiltransferaza (DNMT), ADN-ul celulelor fiice ar rămâne nemetilate, ulterior generând inclusiv demetilare pasivă. DNMT1 este metiltransferaza de întreținere, responsabilă pentru copierea tiparelor de metilare a ADN-ului către catenele fiice în timpul replicării ADN-ului. Șoarecii având ambele copii ale DNMT1 șterse, mor în stadiu embrionar în ziua a 9-a postfertilizare, datorită necesității activității DNMT1 pentru viabilitatea dezvoltării embrionare.[66]
DNMT3a și DNMT3b efectuează metilarea de novo (i.e. noi profiluri de metilare, diferite de ale celulei părinte), având și ele rol esențial în dezvoltarea embrionară.[67] DNMT3L este o proteină omoloagă cu celelalte DNMT3, dar nu are activitate catalitică. În schimb, DNMT3L ajută metiltransferazele omoloage (DNMT3a și DNMT3b), crescându-le capacitatea de legare de ADN și stimulând activitatea lor.
Șoarecii și șobolanii au o a treia enzimă funcțională în metilarea de novo, numită DNMT3C, care a evoluat ca un paralog al Dnmt3b la strămoșul comun al rozătoarelor Muroidea. DNMT3C catalizează metilarea promotorilor elementelor transpozabile în timpul spermatogenezei timpurii, activitate esențială pentru reprimarea lor epigenetică și fertilitatea masculină.[68][69] Nu este încă clar dacă la alte mamifere, care nu au DNMT3C (precum oamenii), DNMT3B sau DNMT3A înlocuiesc funcția acestuia. Un al omolog de ADN metiltransferază identificat este DNMT2 (TRDMT1), care conține toate cele 10 motive comune tuturor ADN-metiltransferazelor; cu toate acestea, DNMT2 (TRDMT1) nu metilează ADN-ul, ci metilează citozina-38 în bucla anticodon a ARN-ului de transfer al acidului aspartic.[70]
Deoarece multe gene supresoare de tumori sunt reprimate prin metilarea ADN-ului în timpul carcinogenezei, au existat încercări de a reactiva aceste gene prin inhibarea DNMT-urilor. 5-Aza-2'-deoxicitidina (decitabina) este un analog nucleozidic care inhibă DNMT-urile prin captarea lor într-un complex covalent pe ADN, ducând la degradarea enzimelor. Cu toate acestea, pentru ca decitabina să fie activă, aceasta trebuie să fie încorporată în genomul celulei, ceea ce poate provoca mutații în celulele fiice, sau moarte celulară. În plus, decitabina este toxică pentru măduva osoasă, ceea ce limitează utilizarea sa terapeutică. Aceste aspecte negative au condus la dezvoltarea de terapii cu ARN antisens care vizează DNMT-urile prin degradarea ARNm-urilor lor și prevenirea traducerii lor.
În plante
S-au făcut progrese semnificative în înțelegerea metilării ADN-ului pentru planta Arabidopsis thaliana. Metilarea ADN-ului la plante diferă de cea de la mamifere: în timp ce metilarea ADN-ului la mamifere are loc în principal pe nucleotida de citozină dintr-un situs CpG, la plante citozina poate fi metilată la situsurile CpG, CpHpG și CpHpH, unde H reprezintă orice nucleotidă, cu excepția guaninei.[71] În general, ADN-ul Arabidopsis este foarte metilat, spectrometriei de masă estimând că 14% din citozine sunt modificabile.[8] Datele de secvențiere a bisulfiților (metodă gold-standard de detectare a metilării de la nivelul ADN-ului) au estimat că aproximativ 25% dintre siturile Arabidopsis CG sunt metilate, dar aceste niveluri variază în funcție de locația geografică (plantele din nord sunt mai mult metilate decât cele sudice).[72]
Principalele ADN-metiltransferaze ale Arabidopsis, sunt: DRM2, MET1 și CMT3. Atât DRM2, cât și MET1, au o omologie semnificativă cu metiltransferazele mamiferelor DNMT3 și DNMT1, în timp ce proteina CMT3 este unică pentru regnul vegetal. DRM2 este singura enzimă implicată în metilarea de novo. Totodată, DRM2, împreună cu MET1 și CMT3, sunt implicate în metilarea de întreținere.[73] Mai sunt și alte ADN-metiltransferaze exprimate în plante, dar încă nu li se cunosc funcțiile (vezi Chromatin Database).
Nivelurile de metilare ale ADN-urilor plantelor variază mult între specii, iar ADN-ul Arabidopsis tinde să fie mai puțin metilate decât cel din alte plante. De exemplu, aproximativ 92,5% din citozinele CpG sunt metilate în Beta vulgaris.[74] De asemenea, modelele de metilare diferă funcție de secvența în care se află citozina; în mod universal, metilarea CpG este mai frecventă decât metilarea CHG și CHH. Metilarea CpG poate fi găsită atât în genele active, cât și în elementele transpozabile, în timp ce metilările CHG și CHH se găsesc de obicei la elementele transpozabile reprimate.[75][71]
Nu este clar modul în care celula determină locațiile metilării ADN-ului de novo, dar dovezile sugerează că pentru multe locații (deși nu toate) este implicată metilarea ADN-ului dirijată de ARN (RdDM). În RdDM, transcrieri de ARN specifice sunt produse dintr-un șablon de ADN genomic, iar acest ARN formează structuri secundare numite molecule de ARN dublu catenar.[76] ARN-urile dublu catenare, fie prin calea ARN-ului mic de interferență (siRNA), fie prin cea a microARN-ului (miARN), direcționează metilarea de novo a locației genomice originare care a produs ARN-ul.[76] Se crede că acest tip de mecanism este important în apărarea celulară împotriva virusurilor ARN și/sau transpozonilor, ambii formând adesea ARN dublu catenare ce pot fi mutagene pentru genomul gazdei. Prin metilarea locațiilor lor genomice, expresia lor este reprimată, protejând genomul de efectul lor mutagen.
La insecte
Diverse ordine de insecte prezintă grade variate de metilare ale ADN-urilor, de la niveluri aproape nedetectabile la muște, la niveluri relativ scăzute la lapidoptere, și mai mari la hemiptere și unii gândaci (până la 14% din toate site-urile CG din Blattella asahinai).[77]
Metilarea funcțională a ADN-ului a fost descoperită la albinele melifere.[78][79] Locurile de metilare ale ADN-ului sunt, în principal, pe corpurile genelor, iar opiniile actuale susțin că aceasta se datorează procesului de reglare a expresiei genelor prin splicing alternativ.[80]
Nivelurile de metilare ale ADN-ului la Drosophila melanogaster sunt aproape nedetectabile.[81] Metode sensibile aplicate la ADN-ul Drosophila sugerează niveluri în intervalul 0,1–0,3% din citozina totală.[82] Un studiu din 2014 a constatat că nivelul scăzut de metilare la musculițele de oțet a apărut „la anumite motive (proteice) scurte, fiind independent de activitatea DNMT2”.[83] În plus, spectrometria de masă[84] a demonstrat prezența unor niveluri foarte scăzute (0,07%), dar semnificative statistic, de metilare ale adeninei în timpul primelor etape ale embriogenezei musculiței de oțet.
În ciuperci
Multe ciuperci au niveluri scăzute (0,1 până la 0,5%) de metilare ale citozinei, în timp ce alte ciuperci au până la 5% din genom metilat.[85] Această valoare pare să varieze atât între specii, cât și în cadrul aceleiași specii.[86] Există, de asemenea, dovezi că metilarea ADN-ului poate fi implicată în controlul specific de status al expresiei genelor la ciuperci. [ <span title="This claim needs references to reliable sources. (November 2010)">citare necesară</span> ] Cu toate acestea, la o limită de detecție de 250 de attomoli, prin utilizarea spectrometriei de masă ultrasenzitive, metilarea ADN-ului nu a fost confirmată la speciile de drojdie unicelulare, cum ar fi Saccharomyces cerevisiae sau Schizosaccharomyces pombe.[8]
Deși drojdiile precum Saccharomyces, Schizosaccharomyces și Aspergillus flavus[87] nu au metilări detectabile ale ADN-urilor, ciuperca filamentoasă Neurospora crassa are un sistem de metilare bine caracterizat.[88] Mai multe gene controlează metilarea în Neurospora și mutația ADN-metiltransferazei, dim-2, scade semnificativ metilările ADN-ului[89], dar nu afectează creșterea sau reproducerea sexuală. În timp ce genomul Neurospora are foarte puțin ADN ce se repetă, jumătate din metilări au loc în ADN-ul recurent, inclusiv relicve de transpozoni și ADN centromeric. Capacitatea de a evalua alte fenomene importante pe un fond genetic cu deficit de ADN-metilază face ca Neurospora să fie un sistem important de studiu al metilării ADN-ului.
În bacterii

Metilarea adeninei sau citozinei este mediată de sistemele de restricție a modificărilor ale multor bacterii, prin care secvențe specifice de ADN sunt metilate periodic în întregul genom.[91] Metilazele sunt enzimele care recunosc secvențele specifice și metilează una dintre bazele din secvența respectivă sau din apropierea ei. ADN-urile străine, nemetilate în acest mod, ce ajung în celulă sunt degradate de enzimele de restricție specifice secvenței. ADN-ul bacterian propriu, nu este recunoscut de aceste enzime de restricție. Acest sistem permite ca bateriile să se autoprotejeze de bacteriofage.
E.coli ADN-adeninmetiltransferaza (Dam) este o enzimă de aproximativ 32 kDa care nu aparține unui sistem de restricție a modificărilor. Secvența specifică de recunoaștere pentru E.coli Dam este GATC, metilarea având loc la poziția N6 a adeninei din această secvență (G meATC). Cele trei perechi de baze care flanchează acest sit influențează, de asemenea, legarea Dam. Dam joacă mai multe roluri cheie în procesele bacteriene, inclusiv repararea nepotrivirilor secvențiale, sincronizarea replicării ADN-ului și expresia genică. Ca rezultat al replicării ADN-ului, starea siturilor GATC din genomul E.coli se schimbă de la complet metilat la semimetilat. Aceasta se datorează faptului că adenina introdusă în noile catene ADN replicate este nemetilată. Remetilarea are loc în decurs de 2-4 secunde, timp în care erorile de replicare din noua catenă sunt reparate. Metilarea, sau absența acesteia, este markerul care permite aparatului de reparare al celulei să diferențieze între șablon și catenele noi replicate. S-a demonstrat că modificarea activității Dam în bacterii are ca rezultat o rată de mutație spontană crescută.
O regiune a ADN-ului care își păstrează starea hemimetilată pentru mai mult timp este originea replicării, care are o abundență de situri GATC. Acest lucru este esențial pentru mecanismul bacterian de sincronizare a replicării ADN-ului. SeqA se leagă de originea replicării, sechestrând-o și prevenind astfel metilarea ei. Deoarece originile hemimetilate ale replicării sunt inactive, acest mecanism limitează replicarea ADN-ului la câte un ciclu celular.
Exprimarea anumitor gene, de exemplu cele care codifică expresia pilusulu în E.coli, este reglată de metilarea situsurilor GATC din regiunea promotor a operonului genei. Condițiile de mediu ale celului, imediat după replicarea ADN-ului determină dacă Dam este blocat de la metilarea unei regiuni proximale sau distale de regiunea promotor. Odată ce profilul de metilare a fost definitivat, transcripția genei pilus este blocată în poziția pornit sau oprit până când celula se divide din nou. La E. coli, acești operoni pili au roluri importante în virulența în infecțiile tractului urinar. S-a propus că inhibitorii Dam pot funcționa ca antibiotice.
Pe de altă parte, ADN-citozinmetilaza (Dcm) vizează situsurile CCAGG și CCTGG pentru a metila citozina la poziția C5 (CmeC(A/T)GG). Cealaltă enzimă metilază, EcoKI, determină metilarea adeninelor în secvențele AAC(N6)GTGC și GCAC(N6)GTT.
În Clostridioides difficile, metilarea ADN-ului la motivul țintă CAAAAA s-a dovedit că are rol în sporulare, un pas cheie în transmiterea bolii, precum și în determinarea lungimii celulei, formarea biofilmului și colonizarea gazdei.[92]
Clonarea moleculară
Majoritatea tulpinilor utilizate de biologii moleculari sunt derivate de E.coli K-12 și posedă atât Dam, cât și Dcm, dar există tulpini disponibile comercial care sunt dam-/dcm- (lipsa de activitate a metilazelor). De fapt, este posibil să se demetileze ADN-ul extras din tulpinile dam+/dcm+, transformându-l în tulpini dam-/dcm-. Acest lucru ar ajuta la digerarea secvențelor care nu sunt recunoscute de enzimele de restricție sensibile la metilare.[93][94]
Enzima de restricție DpnI poate recunoaște siturile 5’-GmeATC-3’ și poate digera ADN-ul metilat. Fiind un motiv atât de scurt, apare frecvent în secvențe din întâmplare și, ca atare, utilizarea sa principală pentru cercetători este de degradare a ADN-ului șablon după PCR (produsele PCR nu sunt metilate datorită lipsei metilazelor din reacție).
Detectare
Metilarea ADN-ului poate fi detectată prin următoarele teste utilizate în prezent în cercetarea științifică:[95]
- Spectometria de masă este o metodă analitică foarte sensibilă și fiabilă pentru detectarea metilării ADN. Totuși, nu oferă informații asupra contextului secvențial de metilare, fiind astfel limitată la studierea funcției acestor modificări asupra ADN-ului.
- PCR specifică metilării (MSP), care se bazează pe reacția bisulfitului de sodiu cu ADN-ul, convertind citozinele nemetilate din CpG, în uracil (UpG), după care se aplică PCR.[96] Totuși, citozinele metilate nu vor fi convertite în acest proces, iar cum primerii sunt concepuți pentru a se atașa siturilor CpG complementare, se poate determina starea respectivei secvențe (metilată sau nemetilată).
- Secvențierea bisulfită a întregului genom, cunoscută și sub denumirea de BS-Seq, este o analiză exhaustivă a metilării ADN-ului întregului genom. Se bazează pe efectul bisulfitului de sodiu asupra ADN-ului genomic, care este apoi secvențiat pe o platformă de Next-generation sequencing. Secvențele obținute sunt apoi realiniate la genomul de referință pentru a determina starea de metilare a dinucleotidelor CpG pe baza nepotrivirilor rezultate datorită conversiei citozinelor nemetilate în uracil.
- Metil-secvențierea enzimatică (EM-seq) funcționează similar cu secvențierea bisulfiților, dar utilizează enzime, APOBEC și TET2 pentru a dezamina citozina nemetilată în uracil. Librăriile EM-seq sunt mai puțin predispuse la deteriorarea ADN-ului decât cele tratate cu bisulfit.[97]
- Secvențierea bisulfită cu reprezentare redusă, cunoscută și sub numele de RRBS, cunoaște mai multe protocoale de lucru. Primul protocol RRBS a fost numit RRBS și vizează aproximativ 10% din metilom, fiind nevoie de un genom de referință. Mai târziu au apărut mai multe protocoale care au putut secvența o porțiune mai mică a genomului și multiplexare mai mare a probei.
- Testul HELP, care se bazează pe capacitatea diferențială a enzimelor de restricție de a recunoaște și scinda situsurile ADN CpG metilate și nemetilate.
- Testul GLAD-PCR, care se bazează pe un nou tip de enzime – endonucleaze ADN sit-specifice, care hidrolizează numai ADN-ul metilat.
- Testele ChIP-on-cip, care se bazează pe capacitatea anticorpilor preparați anterior de a se lega la proteinele asociate metilării ADN-ului, cum ar fi MeCP2.
- Scanarea genomică cu repere de restricție, un test complicat și acum rar utilizat, bazat pe recunoașterea diferențială a enzimelor de restricție a situsurilor CpG metilate și nemetilate; testul este similar ca concept cu testul HELP.
- Imunoprecipitarea ADN-ului metilat (MeDIP), analoagă cu imunoprecipitarea cromatinei, imunoprecipitarea este utilizată pentru a izola fragmente de ADN metilat pentru a fi ulterior utilizate de metodele de identificare a ADN-ului, cum ar fi microarrays-uri ADN (MeDIP-chip) sau secvențierea ADN (MeDIP-seq).
Regiuni metilate diferențiat (DMRs)
Regiunile metilate diferențiat (engl. "differentially methylated regions") sunt regiuni genomice, aparținând diferitelor eșantioane (de țesuturi, celule, indivizi etc.), având stări diferite de metilare și fiind considerate posibile regiuni funcționale implicate în regularea transcripțională a genelor. Identificarea DMR pentru țesuturi multiple (T-DMRs) oferă o perspectivă cuprinzătoare a diferențelor epigenetice existente în țesuturile umane.[98] De exemplu, aceste regiuni metilate diferențiat, unice pentru fiecare țesut în parte, permit diferențierea tipurilor de țesut în laborator (exp. spermă vs. lichid vaginal). Cercetări efectuate de Lee și colab., au arătat că DACT1 și USP49 identifică în mod pozitiv materialul seminal prin examinarea T-DMRs.[99] Utilizarea T-DMRs s-a dovedit utilă în identificarea diferitelor fluide corporale găsite la locul crimei. Cercetătorii din domeniul criminalistic caută noi T-DMRs genomice pentru a le utiliza ca markeri în analiza ADN-ului. Diferența DMRs între eșantioanele canceroase și cele normale demonstrează metilarea aberantă existentă în cancere.[100] Este bine cunoscut faptul că metilarea ADN-ului este asociată cu diferențierea și proliferarea celulelor.[101] Multe DMRs au fost găsite în etapele de dezvoltare (D-DMRs)[102] și în progresul reprogramat (R-DMRs).[103] În plus, există DMRs-uri intra-individuale (Intra-DMRs) ce se corelează cu creșterea vârstei la un anumit individ.[104] Există, de asemenea, DMRs-uri inter-individuale (Inter-DMRs) ce reprezintă modelele diferite de metilare existente între mai mulți indivizi.[105]
QDMR (Quantitative Differentially Methylated Regions) este o abordare cantitativă pentru cuantificarea diferențelor de metilare și identificarea DMRs-urilor din metiloame prin adaptarea entropiei Shannon.[106] Independența de platformă și de specimen a analizei QDMR o face aplicabilă pentru diferite date de metilare. Această abordare oferă un instrument eficient pentru identificarea, cu randament ridicat, a regiunilor funcționale implicate în regularea epigenetică. QDMR poate fi utilizat ca un instrument eficient pentru cuantificarea diferențelor de metilare și pentru identificarea DMRs-urilor din mai multe eșantioane.[107]
Analiza seturilor de gene (i.e. analiza căilor biologice, utilizând instrumente precum DAVID, GoSeq sau GSEA) s-a dovedit a fi sever părtinitoare atunci când este aplicată datelor de metilare de mare capacitate (de exemplu, MeDIP-seq, MeDIP-ChIP, HELP-seq etc. ), și o gamă largă de studii au raportat astfel în mod eronat hipermetilarea genelor legate de dezvoltare și diferențiere; s-a sugerat că acest lucru poate fi corectat folosind permutări ale eșantioanelor sau folosind un model statistic pentru a controla diferențele în numărul de sonde/situsuri CpG care vizează fiecare genă.[108]
Fiecare regiune metilată diferențial este caracterizată printr-o serie de markeri de metilare ai ADN-ului (engl. "DNA methylation marks"). Deci, pe când markerii de metilare ai ADN-ului se referă la metilările efective de la nivelul nucleotidelor, regiunile metilate diferențial se referă la profilurile de metilare pentru secvențe de ADN (fiecare profil fiind caracterizat de o anumită dispunere a markerilor de metilare).
Instrumentul de analiză și raportare a metilării specifice bazat pe entropie, denumit „SMART”, se concentrează pe integrarea unui număr mare de metilomi ADN pentru identificarea de novo a semnelor de metilare specifice tipului de celule. Cea mai recentă versiune a SMART are trei funcții principale, inclusiv identificarea de novo a regiunilor metilate diferențial (DMRs) prin segmentarea genomului, identificarea DMRs-urilor din anumite regiuni de interes și identificarea siturilor CpG metilate diferențial.[109]
În identificarea și detectarea fluidelor corporale
Metilarea ADN-ului permite identificarea unor cantități mici de fluid corporal prin utilizarea ADN-ului extras. De obicei, cele două abordări utilizate pentru acest scop sunt: metoda enzimelor restrictive sensibile la metilare și tratamentul cu bisulfit de sodiu.[110] Enzimele restrictive sensibile la metilare funcționează prin scindarea perechilor CpG metilate. Incidental, citozinele nemetilate sunt transformate în uracil. În special, profilurile de metilare pot oferi informații despre când și cum au fost lăsate fluidele corporale la locul crimei, pot identifica tipul de fluid corporal și pot aproxima vârsta, sexul și caracteristicile fenotipice ale făptuitorilor.[111] Decizia markerilor de metilare utilizați reprezintă primul pas pentru identificarea fluidelor corporale. Profilurile de metilare țesut-specifice se bazează pe cercetări anterioare. Gradul de metilare pentru regiunile metilate diferențiat țesut-specifice (engl. "T-DMRs") variază în funcție de fluidul corporal.[111] Câteva exemple de markeri de metilare ADN sunt: Mens1 (sânge menstrual), Spei1 (salivă) și Sperm2 (lichidul seminal).
Metilarea ADN-ului oferă o bună sensibilitate la identificarea și detectarea fluidelor corporale. Într-un studiu, doar zece nanograme dintr-un eșantion au fost necesare pentru a stabili rezultate de succes.[112] Metilarea ADN-ului oferă capacitatea de a discerne în cazul eșantioanelor mixte. Metilarea ADN-ului nu este insensibilă la condițiile externe, însă chiar și în condiții nefavorabile, markerii de metilare sunt suficient de stabili pentru a permite identificarea eșantionului. Într-un studiu s-a constatat că nu există modificări semnificative în modelele de metilare ale eșantioanelor datorate trecerii timpului.[111]
Detectarea metilării ADN-ului din fluide a devenit una dintre principalele abordări ale biopsiei de lichide.[113] Utilizarea algoritmilor cu semnale de deconvoluție asupra eșantioanelor ADN din fluide permite localizarea cancerelor primare, a rejecției de alogrefă și detectarea rezistenței la terapia hormonală.[114]
Note
- ^ D. B. Dunn, J. D. Smith: "The occurrence of 6-methylaminopurine in deoxyribonucleic acids". In: Biochem J. 68(4), Apr 1958, S. 627–636. PMID 13522672. PMC 1200409.
- ^ Melanie Ehrlich, Miguel A. Gama-Sosa, Laura H. Carreira, Lars G. Ljungdahl, Kenneth C. Kuo, Charles W. Gehrke: "DNA methylation in thermophilic bacteria: N6-methylcytosine, 5-methylcytosine, and N6-methyladenine." In: Nucleic Acids Research. 13, 1985, S. 1399. PMID 4000939. PMC 341080.
- ^ „Methylation of the deoxyribonucleic acid of Physarum polycephalum at various periods during the mitotic cycle”. The Journal of Biological Chemistry. 245 (23): 6436–6441. decembrie 1970. doi:10.1016/S0021-9258(18)62627-4. PMID 5530731.

- ^ „Lack of 5-methylcytosine in Dictyostelium discoideum DNA”. The Biochemical Journal. 277 (1): 273–275. iulie 1991. doi:10.1042/bj2770273. PMC 1151219
 . PMID 1713034.
. PMID 1713034. - ^ „The Dictyostelium discoideum genome lacks significant DNA methylation and uncovers palindromic sequences as a source of false positives in bisulfite sequencing”. NAR Genomics Bioinformatics. 5 (2): lqad035. aprilie 2023. doi:10.1093/nargab/lqad035. PMC 10111430 . PMID 37081864 Verificați valoarea
|pmid=(ajutor). - ^ „Trace analysis of methylated and hydroxymethylated cytosines in DNA by isotope-dilution LC-MS/MS: first evidence of DNA methylation in Caenorhabditis elegans”. The Biochemical Journal. 465 (1): 39–47. ianuarie 2015. doi:10.1042/bj20140844. PMID 25299492.
- ^ „Molecular biology. Methylation talk between histones and DNA”. Science. 294 (5549): 2113–2115. decembrie 2001. doi:10.1126/science.1066726. PMID 11739943.
As a result of this process, known as repeat-induced point mutation (RIP), the wild-type Neurospora genome contains a small fraction of methylated DNA, the majority of the DNA remaining nonmethylated.
- ^ a b c „Cytosine DNA methylation is found in Drosophila melanogaster but absent in Saccharomyces cerevisiae, Schizosaccharomyces pombe, and other yeast species”. Analytical Chemistry. 86 (8): 3697–3702. aprilie 2014. doi:10.1021/ac500447w. PMC 4006885 . PMID 24640988.
- ^ „N6-methyladenine: the other methylated base of DNA”. BioEssays. 28 (3): 309–315. martie 2006. doi:10.1002/bies.20342. PMC 2754416 . PMID 16479578.
- ^ „DNA methylation on N(6)-adenine in mammalian embryonic stem cells”. Nature. 532 (7599): 329–333. aprilie 2016. Bibcode:2016Natur.532..329W. doi:10.1038/nature17640. PMC 4977844 . PMID 27027282. Mentenanță CS1: display-autori (link)
- ^ Angéla Békési and Beáta G Vértessy "Uracil in DNA: error or signal?"
- ^ „Reviving the RNA World: An Insight into the Appearance of RNA Methyltransferases”. Frontiers in Genetics. 7: 99. . doi:10.3389/fgene.2016.00099. PMC 4893491 . PMID 27375676.
- ^ „De novo methylation of MMLV provirus in embryonic stem cells: CpG versus non-CpG methylation”. Gene. 289 (1–2): 41–48. mai 2002. doi:10.1016/S0378-1119(02)00469-9. PMID 12036582.
- ^ „Allele-specific non-CpG methylation of the Nf1 gene during early mouse development”. Developmental Biology. 240 (2): 585–598. decembrie 2001. doi:10.1006/dbio.2001.0504. PMID 11784085.
- ^ a b „Human DNA methylomes at base resolution show widespread epigenomic differences”. Nature. 462 (7271): 315–322. noiembrie 2009. Bibcode:2009Natur.462..315L. doi:10.1038/nature08514. PMC 2857523 . PMID 19829295. Mentenanță CS1: display-autori (link)
- ^ „Global epigenomic reconfiguration during mammalian brain development”. Science. 341 (6146): 1237905. august 2013. doi:10.1126/science.1237905. PMC 3785061 . PMID 23828890. Mentenanță CS1: display-autori (link)
- ^ a b „Whole-genome fingerprint of the DNA methylome during human B cell differentiation”. Nature Genetics. 47 (7): 746–756. iulie 2015. doi:10.1038/ng.3291. PMC 5444519 . PMID 26053498. Mentenanță CS1: display-autori (link)
- ^ „DNA methylation: an introduction to the biology and the disease-associated changes of a promising biomarker”. Molecular Biotechnology. 44 (1): 71–81. ianuarie 2010. doi:10.1007/s12033-009-9216-2. PMID 19842073.
- ^ „DNA-binding factors shape the mouse methylome at distal regulatory regions”. Nature. 480 (7378): 490–495. decembrie 2011. doi:10.1038/nature11086. PMID 22170606. Mentenanță CS1: display-autori (link)
- ^ a b c „Genome-wide evolutionary analysis of eukaryotic DNA methylation”. Science. 328 (5980): 916–919. mai 2010. Bibcode:2010Sci...328..916Z. doi:10.1126/science.1186366. PMID 20395474.
Here we quantify DNA methylation in seventeen eukaryotic genomes....
Supplemental figures appear to be only accessible via the science.sciencemag.org paywall. - ^ „CpG methylation is targeted to transcription units in an invertebrate genome”. Genome Research. 17 (5): 625–631. mai 2007. doi:10.1101/gr.6163007. PMC 1855171 . PMID 17420183.
- ^ a b „Initial sequencing and analysis of the human genome”. Nature. 409 (6822): 860–921. februarie 2001. Bibcode:2001Natur.409..860L. doi:10.1038/35057062. PMID 11237011. Mentenanță CS1: display-autori (link)
- ^ „CpG-rich islands and the function of DNA methylation”. Nature. 321 (6067): 209–213. . Bibcode:1986Natur.321..209B. doi:10.1038/321209a0. PMID 2423876.
- ^ „CpG islands in vertebrate genomes”. Journal of Molecular Biology. 196 (2): 261–282. iulie 1987. doi:10.1016/0022-2836(87)90689-9. PMID 3656447.
- ^ „Orphan CpG islands identify numerous conserved promoters in the mammalian genome”. PLOS Genetics. 6 (9): e1001134. septembrie 2010. doi:10.1371/journal.pgen.1001134. PMC 2944787 . PMID 20885785. Mentenanță CS1: display-autori (link)
- ^ „A genome-wide analysis of CpG dinucleotides in the human genome distinguishes two distinct classes of promoters”. Proceedings of the National Academy of Sciences of the United States of America. 103 (5): 1412–1417. ianuarie 2006. Bibcode:2006PNAS..103.1412S. doi:10.1073/pnas.0510310103. PMC 1345710 . PMID 16432200.
- ^ a b „Conservation and divergence of methylation patterning in plants and animals”. Proceedings of the National Academy of Sciences of the United States of America. 107 (19): 8689–8694. mai 2010. doi:10.1073/pnas.1002720107. PMC 2889301 . PMID 20395551. Mentenanță CS1: display-autori (link)
- ^ „Lineage-specific polycomb targets and de novo DNA methylation define restriction and potential of neuronal progenitors”. Molecular Cell. 30 (6): 755–766. iunie 2008. doi:10.1016/j.molcel.2008.05.007. PMID 18514006. Mentenanță CS1: display-autori (link)
- ^ „Distribution, silencing potential and evolutionary impact of promoter DNA methylation in the human genome”. Nature Genetics. 39 (4): 457–466. aprilie 2007. doi:10.1038/ng1990. PMID 17334365.
- ^ „Function and information content of DNA methylation”. Nature. 517 (7534): 321–326. ianuarie 2015. Bibcode:2015Natur.517..321S. doi:10.1038/nature14192. PMID 25592537.
- ^ „Genome-wide conserved consensus transcription factor binding motifs are hyper-methylated”. BMC Genomics. 11 (1): 519. septembrie 2010. doi:10.1186/1471-2164-11-519. PMC 2997012 . PMID 20875111.
- ^ „Genome-wide analysis in the mouse embryo reveals the importance of DNA methylation for transcription integrity”. Nature Communications. 11 (1): 3153. iunie 2020. Bibcode:2020NatCo..11.3153D. doi:10.1038/s41467-020-16919-w. PMC 7305168 . PMID 32561758. Mentenanță CS1: display-autori (link)
- ^ „Dnmt1-independent CG methylation contributes to nucleosome positioning in diverse eukaryotes”. Cell. 156 (6): 1286–1297. martie 2014. doi:10.1016/j.cell.2014.01.029. PMC 3969382 . PMID 24630728.
- ^ „Cytosine methylation and the ecology of intragenomic parasites”. Trends in Genetics. 13 (8): 335–340. august 1997. doi:10.1016/s0168-9525(97)01181-5. PMID 9260521.
- ^ „DNA methylation enables transposable element-driven genome expansion”. Proceedings of the National Academy of Sciences of the United States of America. 117 (32): 19359–19366. august 2020. Bibcode:2020PNAS..11719359Z. doi:10.1073/pnas.1921719117. PMC 7431005 . PMID 32719115.
- ^ „The alternative role of DNA methylation in splicing regulation”. Trends in Genetics. 31 (5): 274–280. mai 2015. doi:10.1016/j.tig.2015.03.002. PMID 25837375.
- ^ „Conserved role of intragenic DNA methylation in regulating alternative promoters”. Nature. 466 (7303): 253–257. iulie 2010. Bibcode:2010Natur.466..253M. doi:10.1038/nature09165. PMC 3998662 . PMID 20613842. Mentenanță CS1: display-autori (link)
- ^ „Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription”. Cell. 123 (4): 581–592. noiembrie 2005. doi:10.1016/j.cell.2005.10.023. PMID 16286007. Mentenanță CS1: display-autori (link)
- ^ a b „Global DNA methylation and chondrogenesis of rat limb buds in a three-dimensional organ culture system”. Bosnian Journal of Basic Medical Sciences. 22 (4): 560–568. iulie 2022. doi:10.17305/bjbms.2021.6584. PMC 9392980 . PMID 35188093 Verificați valoarea
|pmid=(ajutor). Mentenanță CS1: display-autori (link) - ^ „Loss of methylation activates Xist in somatic but not in embryonic cells”. Genes & Development. 9 (19): 2325–2334. octombrie 1995. doi:10.1101/gad.9.19.2325. PMID 7557385.
- ^ „Role for DNA methylation in genomic imprinting”. Nature. 366 (6453): 362–365. noiembrie 1993. Bibcode:1993Natur.366..362L. doi:10.1038/366362a0. PMID 8247133.
- ^ „Reprogramming DNA methylation in the mammalian life cycle: building and breaking epigenetic barriers”. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 368 (1609): 20110330. ianuarie 2013. doi:10.1098/rstb.2011.0330. PMC 3539359 . PMID 23166394.
- ^ „Metabolic recoding of epigenetics in cancer”. Cancer Communications. 38 (1): 25. mai 2018. doi:10.1186/s40880-018-0302-3. PMC 5993135 . PMID 29784032.
- ^ „Specific gene hypomethylation and cancer: new insights into coding region feature trends”. Bioinformation. 3 (8): 340–343. aprilie 2009. doi:10.6026/97320630003340. PMC 2720671 . PMID 19707296.
- ^ Epigenetics: A Reference Manual. Caister Academic Press. . ISBN 978-1-904455-88-2.
- ^ a b c „Epigenetic alterations in aging”. Journal of Applied Physiology. 109 (2): 586–597. august 2010. doi:10.1152/japplphysiol.00238.2010. PMC 2928596 . PMID 20448029.
- ^ „DNA methylation polymorphisms precede any histological sign of atherosclerosis in mice lacking apolipoprotein E”. The Journal of Biological Chemistry. 279 (28): 29147–29154. iulie 2004. doi:10.1074/jbc.m403618200. PMID 15131116. Mentenanță CS1: display-autori (link)
- ^ „Increased homocysteine and S-adenosylhomocysteine concentrations and DNA hypomethylation in vascular disease”. Clinical Chemistry. 49 (8): 1292–1296. august 2003. doi:10.1373/49.8.1292. PMID 12881445. Mentenanță CS1: display-autori (link)
- ^ „Hypermethylation of estrogen receptor-alpha gene in atheromatosis patients and its correlation with homocysteine”. Pathophysiology. 16 (4): 259–265. octombrie 2009. doi:10.1016/j.pathophys.2009.02.010. PMID 19285843.
- ^ „DNA methylation and atherosclerosis”. The Journal of Nutrition. 132 (8 Suppl): 2406S–2409S. august 2002. doi:10.1093/jn/132.8.2406S. PMID 12163701.
- ^ „Methylation of the estrogen receptor-alpha gene promoter is selectively increased in proliferating human aortic smooth muscle cells”. Cardiovascular Research. 46 (1): 172–179. aprilie 2000. doi:10.1016/s0008-6363(00)00004-3. PMID 10727665. Mentenanță CS1: display-autori (link)
- ^ „Inactivation of monocarboxylate transporter MCT3 by DNA methylation in atherosclerosis”. Circulation. 112 (9): 1353–1361. august 2005. doi:10.1161/circulationaha.104.519025. PMID 16116050.
- ^ „Genome-wide DNA methylation encodes cardiac transcriptional reprogramming in human ischemic heart failure”. Laboratory Investigation; A Journal of Technical Methods and Pathology. 99 (3): 371–386. martie 2019. doi:10.1038/s41374-018-0104-x. PMC 6515060 . PMID 30089854. Mentenanță CS1: display-autori (link)
- ^ „Racial and socioeconomic disparity associates with differences in cardiac DNA methylation among men with end-stage heart failure”. American Journal of Physiology. Heart and Circulatory Physiology. 320 (5): H2066–H2079. mai 2021. doi:10.1152/ajpheart.00036.2021. PMC 8163657 . PMID 33769919. Mentenanță CS1: display-autori (link)
- ^ „A longitudinal study of epigenetic variation in twins”. Epigenetics. 5 (6): 516–526. august 2010. doi:10.4161/epi.5.6.12226. PMC 3322496 . PMID 20505345. Mentenanță CS1: display-autori (link)
- ^ a b „Distinct DNA methylomes of newborns and centenarians”. Proceedings of the National Academy of Sciences of the United States of America. 109 (26): 10522–10527. iunie 2012. Bibcode:2012PNAS..10910522H. doi:10.1073/pnas.1120658109. PMC 3387108 . PMID 22689993. Mentenanță CS1: display-autori (link)
- ^ a b „Acute exercise remodels promoter methylation in human skeletal muscle”. Cell Metabolism. 15 (3): 405–411. martie 2012. doi:10.1016/j.cmet.2012.01.001. PMID 22405075. Mentenanță CS1: display-autori (link)
- ^ „A six months exercise intervention influences the genome-wide DNA methylation pattern in human adipose tissue”. PLOS Genetics. 9 (6): e1003572. iunie 2013. doi:10.1371/journal.pgen.1003572. PMC 3694844 . PMID 23825961. Mentenanță CS1: display-autori (link)
- ^ a b „Dynamic DNA methylation controls glutamate receptor trafficking and synaptic scaling”. Journal of Neurochemistry. 137 (3): 312–330. mai 2016. doi:10.1111/jnc.13564. PMC 4836967 . PMID 26849493.
- ^ „Epigenetic regulation and chromatin remodeling in learning and memory”. Experimental & Molecular Medicine. 49 (1): e281. ianuarie 2017. doi:10.1038/emm.2016.140. PMC 5291841 . PMID 28082740.
- ^ „Memory consolidation for contextual and auditory fear conditioning is dependent on protein synthesis, PKA, and MAP kinase”. Learning & Memory. 6 (2): 97–110. . doi:10.1101/lm.6.2.97. PMC 311283 . PMID 10327235.
- ^ a b „DNA methylation changes in plasticity genes accompany the formation and maintenance of memory”. Nature Neuroscience. 19 (1): 102–110. ianuarie 2016. doi:10.1038/nn.4194. PMC 4700510 . PMID 26656643. Mentenanță CS1: display-autori (link)
- ^ a b „Experience-dependent epigenomic reorganization in the hippocampus”. Learning & Memory. 24 (7): 278–288. iulie 2017. doi:10.1101/lm.045112.117. PMC 5473107 . PMID 28620075.
- ^ a b „DNA Methylation and Establishing Memory”. Epigenetics Insights. 15: 25168657211072499. . doi:10.1177/25168657211072499. PMC 8793415 . PMID 35098021 Verificați valoarea
|pmid=(ajutor). - ^ „Review on DNA Methylation”. METHODS.info.
- ^ Jin, Bilian; Robertson, Keith D. (), Karpf, Adam R., ed., „DNA Methyltransferases, DNA Damage Repair, and Cancer”, Epigenetic Alterations in Oncogenesis (în engleză), Springer New York, 754, pp. 3–29, doi:10.1007/978-1-4419-9967-2_1, ISBN 978-1-4419-9966-5, PMC 3707278 , PMID 22956494, accesat în Mentenanță CS1: Format PMC (link)
- ^ Jin, Bilian; Robertson, Keith D. (), Karpf, Adam R., ed., „DNA Methyltransferases, DNA Damage Repair, and Cancer”, Epigenetic Alterations in Oncogenesis (în engleză), Springer New York, 754, pp. 3–29, doi:10.1007/978-1-4419-9967-2_1, ISBN 978-1-4419-9966-5, PMC 3707278 , PMID 22956494, accesat în Mentenanță CS1: Format PMC (link)
- ^ „The DNA methyltransferase DNMT3C protects male germ cells from transposon activity”. Science. 354 (6314): 909–912. noiembrie 2016. Bibcode:2016Sci...354..909B. doi:10.1126/science.aah5143. PMID 27856912. Mentenanță CS1: display-autori (link)
- ^ „rahu is a mutant allele of Dnmt3c, encoding a DNA methyltransferase homolog required for meiosis and transposon repression in the mouse male germline”. PLOS Genetics. 13 (8): e1006964. august 2017. doi:10.1371/journal.pgen.1006964. PMC 5607212 . PMID 28854222. Mentenanță CS1: display-autori (link)
- ^ „Methylation of tRNAAsp by the DNA methyltransferase homolog Dnmt2”. Science. 311 (5759): 395–398. ianuarie 2006. Bibcode:2006Sci...311..395G. doi:10.1126/science.1120976. PMID 16424344. Mentenanță CS1: display-autori (link)
- ^ a b „Dynamics and function of DNA methylation in plants”. Nature Reviews. Molecular Cell Biology. 19 (8): 489–506. august 2018. doi:10.1038/s41580-018-0016-z. PMID 29784956.
- ^ „DNA methylation in Arabidopsis has a genetic basis and shows evidence of local adaptation”. eLife. 4: e05255. mai 2015. doi:10.7554/eLife.05255. PMC 4413256 . PMID 25939354. Mentenanță CS1: display-autori (link)
- ^ „Locus-specific control of asymmetric and CpNpG methylation by the DRM and CMT3 methyltransferase genes”. Proceedings of the National Academy of Sciences of the United States of America. 99 (Suppl 4): 16491–16498. decembrie 2002. Bibcode:2002PNAS...9916491C. doi:10.1073/pnas.162371599. PMC 139913 . PMID 12151602.
- ^ „Widespread natural variation of DNA methylation within angiosperms”. Genome Biology. 17 (1): 194. septembrie 2016. doi:10.1186/s13059-016-1059-0. PMC 5037628 . PMID 27671052. Mentenanță CS1: display-autori (link)
- ^ „Genome-wide evolutionary analysis of eukaryotic DNA methylation”. Science. 328 (5980): 916–919. mai 2010. Bibcode:2010Sci...328..916Z. doi:10.1126/science.1186366. PMID 20395474.
- ^ a b „RNA-directed DNA methylation in Arabidopsis”. Proceedings of the National Academy of Sciences of the United States of America. 99 (Suppl 4): 16499–16506. decembrie 2002. Bibcode:2002PNAS...9916499A. doi:10.1073/pnas.162371499. PMC 139914 . PMID 12169664.
- ^ „Evolution of DNA Methylation across Insects”. Molecular Biology and Evolution. 34 (3): 654–665. martie 2017. doi:10.1093/molbev/msw264. PMC 5400375 . PMID 28025279.
- ^ „Functional CpG methylation system in a social insect”. Science. 314 (5799): 645–647. octombrie 2006. Bibcode:2006Sci...314..645W. doi:10.1126/science.1135213. PMID 17068262. Mentenanță CS1: display-autori (link)
- ^ Ying and Li-Byarlay (). Physiological and Molecular Mechanisms of Nutrition in Honey Bees. Advances in Insect Physiology. 49. pp. 25–58. doi:10.1016/bs.aiip.2015.06.002. ISBN 9780128025864.
- ^ „RNA interference knockdown of DNA methyl-transferase 3 affects gene alternative splicing in the honey bee”. Proceedings of the National Academy of Sciences of the United States of America. 110 (31): 12750–12755. iulie 2013. Bibcode:2013PNAS..11012750L. doi:10.1073/pnas.1310735110. PMC 3732956 . PMID 23852726. Mentenanță CS1: display-autori (link)
- ^ „The two-dimensional restriction analysis of Drosophila DNAs: males and females”. Gene. 13 (4): 395–408. mai 1981. doi:10.1016/0378-1119(81)90019-6. PMID 6266924.
- ^ „DNA methylation in Drosophila melanogaster”. Nature. 408 (6812): 538–540. noiembrie 2000. doi:10.1038/35046205. PMID 11117732.
- ^ „Genome methylation in D. melanogaster is found at specific short motifs and is independent of DNMT2 activity”. Genome Research. 24 (5): 821–830. mai 2014. doi:10.1101/gr.162412.113. PMC 4009611 . PMID 24558263. Mentenanță CS1: display-autori (link)
- ^ „N6-methyladenine DNA modification in Drosophila”. Cell. 161 (4): 893–906. mai 2015. doi:10.1016/j.cell.2015.04.018. PMID 25936838. Mentenanță CS1: display-autori (link)
- ^ „DNA methylation in the fungi”. The Journal of Biological Chemistry. 259 (13): 8033–8036. iulie 1984. doi:10.1016/S0021-9258(17)39681-3. PMID 6330093.
- ^ „A comparison of DNA methylation levels in selected isolates of higher fungi”. Mycologia. 90 (5): 785–790. . doi:10.2307/3761319. JSTOR 3761319.
- ^ „Bisulfite sequencing reveals that Aspergillus flavus holds a hollow in DNA methylation”. PLOS ONE. 7 (1): e30349. . Bibcode:2012PLoSO...730349L. doi:10.1371/journal.pone.0030349. PMC 3262820 . PMID 22276181. Mentenanță CS1: display-autori (link)
- ^ „The methylated component of the Neurospora crassa genome”. Nature. 422 (6934): 893–897. aprilie 2003. Bibcode:2003Natur.422..893S. doi:10.1038/nature01564. PMID 12712205.
- ^ Möller, Mareike; Habig, Michael; Lorrain, Cécile; Feurtey, Alice; Haueisen, Janine; Fagundes, Wagner C.; Alizadeh, Alireza; Freitag, Michael; Stukenbrock, Eva H. (), „Recent loss of the Dim2 DNA methyltransferase decreases mutation rate in repeats and changes evolutionary trajectory in a fungal pathogen”, PLoS Genetics, 17 (3), pp. e1009448, doi:10.1371/journal.pgen.1009448, ISSN 1553-7390, PMC 8016269 , PMID 33750960, accesat în
- ^ Matthew J. Blow, Tyson A. Clark, Chris G. Daum, Adam M. Deutschbauer, Alexey Fomenkov, Roxanne Fries, Jeff Froula, Dongwan D. Kang, Rex R. Malmstrom, Richard D. Morgan, Janos Posfai, Kanwar Singh, Axel Visel, Kelly Wetmore, Zhiying Zhao, Edward M. Rubin, Jonas Korlach, Len A. Pennacchio, Richard J. Roberts: The Epigenomic Landscape of Prokaryotes. In: PLoS Genet. 12(2), Feb 2016, S. e1005854. doi:10.1371/journal.pgen.1005854. PMID 26870957. PMC 4752239
- ^ „Bacterial Epigenomics: Coming of Age”. mSystems. 6 (4): e0074721. august 2021. doi:10.1128/mSystems.00747-21. PMC 8407109 . PMID 34402642.
- ^ „Epigenomic characterization of Clostridioides difficile finds a conserved DNA methyltransferase that mediates sporulation and pathogenesis”. Nature Microbiology. 5 (1): 166–180. ianuarie 2020. doi:10.1038/s41564-019-0613-4. PMC 6925328 . PMID 31768029. Mentenanță CS1: display-autori (link)
- ^ „The dam and dcm strains of Escherichia coli--a review”. Gene. 143 (1): 1–12. mai 1994. doi:10.1016/0378-1119(94)90597-5. PMID 8200522.
- ^ „Making unmethylated (dam-/dcm-) DNA”. Arhivat din original la .
- ^ „Crime investigation through DNA methylation analysis: methods and applications in forensics”. Egyptian Journal of Forensic Sciences. 8 (7). ianuarie 2018. doi:10.1186/s41935-018-0042-1.
- ^ „Optimizing methodologies for PCR-based DNA methylation analysis”. BioTechniques. 55 (4): 181–197. octombrie 2013. doi:10.2144/000114087. PMID 24107250.
- ^ Feng, Suhua; Zhong, Zhenhui; Wang, Ming; Jacobsen, Steven E. (). „Efficient and accurate determination of genome-wide DNA methylation patterns in Arabidopsis thaliana with enzymatic methyl sequencing”. Epigenetics & Chromatin. 13 (1): 42. doi:10.1186/s13072-020-00361-9. ISSN 1756-8935. PMC 7542392 . PMID 33028374.
- ^ „An integrated resource for genome-wide identification and analysis of human tissue-specific differentially methylated regions (tDMRs)”. Genome Research. 18 (9): 1518–1529. septembrie 2008. doi:10.1101/gr.077479.108. PMC 2527707 . PMID 18577705. Mentenanță CS1: display-autori (link)
- ^ „Potential forensic application of DNA methylation profiling to body fluid identification”. International Journal of Legal Medicine. 126 (1): 55–62. ianuarie 2012. doi:10.1007/s00414-011-0569-2. PMID 21626087.
- ^ „The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores”. Nature Genetics. 41 (2): 178–186. februarie 2009. doi:10.1038/ng.298. PMC 2729128 . PMID 19151715. Mentenanță CS1: display-autori (link)
- ^ „Epigenetic reprogramming in mammalian development”. Science. 293 (5532): 1089–1093. august 2001. doi:10.1126/science.1063443. PMID 11498579.
- ^ „Genome-scale DNA methylation maps of pluripotent and differentiated cells”. Nature. 454 (7205): 766–770. august 2008. Bibcode:2008Natur.454..766M. doi:10.1038/nature07107. PMC 2896277 . PMID 18600261. Mentenanță CS1: display-autori (link)
- ^ „Differential methylation of tissue- and cancer-specific CpG island shores distinguishes human induced pluripotent stem cells, embryonic stem cells and fibroblasts”. Nature Genetics. 41 (12): 1350–1353. decembrie 2009. doi:10.1038/ng.471. PMC 2958040 . PMID 19881528. Mentenanță CS1: display-autori (link)
- ^ „Intra-individual change over time in DNA methylation with familial clustering”. JAMA. 299 (24): 2877–2883. iunie 2008. doi:10.1001/jama.299.24.2877. PMC 2581898 . PMID 18577732. Mentenanță CS1: display-autori (link)
- ^ „Inter-individual variation of DNA methylation and its implications for large-scale epigenome mapping”. Nucleic Acids Research. 36 (10): e55. iunie 2008. doi:10.1093/nar/gkn122. PMC 2425484 . PMID 18413340.
- ^ „QDMR: a quantitative method for identification of differentially methylated regions by entropy”. bioinfo.hrbmu.edu.cn. Arhivat din original la . Accesat în .
- ^ „QDMR: a quantitative method for identification of differentially methylated regions by entropy”. Nucleic Acids Research. 39 (9): e58. mai 2011. doi:10.1093/nar/gkr053. PMC 3089487 . PMID 21306990. Mentenanță CS1: display-autori (link)
- ^ „Gene-set analysis is severely biased when applied to genome-wide methylation data”. Bioinformatics. 29 (15): 1851–1857. august 2013. doi:10.1093/bioinformatics/btt311. PMID 23732277.
- ^ „SMART 2: A Comprehensive Analysis Tool for Bisulfite Sequencing Data”. fame.edbc.org. .
- ^ „Molecular approaches for forensic cell type identification: On mRNA, miRNA, DNA methylation and microbial markers”. Forensic Science International. Genetics. 18: 21–32. septembrie 2015. doi:10.1016/j.fsigen.2014.11.015. PMID 25488609.
- ^ a b c „DNA methylation and application in forensic sciences”. Forensic Science International. 249: 255–265. aprilie 2015. doi:10.1016/j.forsciint.2015.01.037. PMID 25732744. Eroare la citare: Etichetă
<ref>invalidă; numele ":4" este definit de mai multe ori cu conținut diferit - ^ „Developmental validation studies of epigenetic DNA methylation markers for the detection of blood, semen and saliva samples”. Forensic Science International. Genetics. 23: 55–63. iulie 2016. doi:10.1016/j.fsigen.2016.01.017. PMID 27010659.
- ^ „Principles of DNA methylation and their implications for biology and medicine”. Lancet. 392 (10149): 777–786. septembrie 2018. doi:10.1016/S0140-6736(18)31268-6. PMID 30100054.
- ^ „Comprehensive human cell-type methylation atlas reveals origins of circulating cell-free DNA in health and disease”. Nature Communications. 9 (1): 5068. noiembrie 2018. Bibcode:2018NatCo...9.5068M. doi:10.1038/s41467-018-07466-6. PMC 6265251 . PMID 30498206. Mentenanță CS1: display-autori (link)

Vezi și
- Epigenetica, la care metilarea ADN-ului contribuie semnificativ
- Ceas epigenetic, o metodă de calcul a vârstei subiecților bazată pe metilarea ADN-ului
- Genomul
- Imprimarea genomică, ce reprezintă reprimări moștenite ale alelelor realizate cu ajutorul metilării ADN-ului
Legături externe
- DNA Methylation la Medical Subject Headings (MeSH), de la Biblioteca Națională de Medicină din Statele Unite
- ENCODE threads explorer Non-coding RNA characterization. Nature (journal)
- PCMdb Pancreatic Cancer Methylation Database.
- SMART Specific Methylation Analysis and Report Tool
- Human Methylation Mark Atlas
- DiseaseMeth Arhivat în , la Wayback Machine. Human disease methylation database
- EWAS Atlas A knowledgebase of epigenome-wide association studies










