ケレンケン
| ケレンケン | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 マウント | |||||||||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||||||||
| 中新世中期 | |||||||||||||||||||||||||||
| 分類 | |||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
| 学名 | |||||||||||||||||||||||||||
| Kelenken Bertelli et al., 2007 | |||||||||||||||||||||||||||
| タイプ種 | |||||||||||||||||||||||||||
| Kelenken guillermoi Bertelli et al., 2007 |
ケレンケン(学名:Kelenken)は、約1500万年前にあたる新第三紀の中期中新世(英語版)に現在でいうアルゼンチンに生息した、絶滅した大型肉食鳥類フォルスラコス科の属。唯一既知の標本はパタゴニア地域のコマロ(英語版)で当時の高校生Guillermo Aguirre-Zabalaにより発見され、2007年に新属新種Kelenken guillermoiのホロタイプ標本に指定された。属名はテウェルチェ族の神話に登場する霊に由来し、種小名は発見者への献名である。ホロタイプ標本は既知の大型フォルスラコス科の中で最も完全な頭蓋骨の1つから構成されており、また足根中足骨(英語版)や趾骨を含む。ケレンケンの発見により、従来理解が乏しかった部位について大型フォルスラコス科の解剖学的特徴の理解が明確化した。分類体系としては、デヴィケンジア(英語版)やフォルスラコスと共にフォルスラコス亜科に属する。
発見と命名


左側から見たホロタイプ標本の頭蓋骨
2004年頃、アルゼンチンのリオネグロ州北西部の小さな村であるコマロ(英語版)の鉄道から約100メートル離れた地点(南緯41度01分59.4秒 西経70度15分29.7秒 / 南緯41.033167度 西経70.258250度 / -41.033167; -70.258250)にて、2軒の家屋の間でアルゼンチンの高校生Guillermo Aguirre-Zabalaがフォルスラコス科の鳥類の化石を発見した。標本が発見された露頭はCollón Curá層のものである。Aguirre-Zabalaは自身で化石のプレパレーションを行い、この化石の発見を切っ掛けとして、心理学から古生物学と地球科学へ志を転換した[1][2][3]。
標本はリオネグロ州のMuseo Asociación Paleontológica Barilocheのコレクションに加えられ、BAR 3877-11という標本番号が付与された[3]。この動物の命名以前に、アルゼンチンの古生物学者ルイス・キアッペとサラ・ベルテッリは2006年の短報で当該標本を報告・議論した[3][4]。Bertelli (2007)は本標本をホロタイプ標本としてKelenken guillermoiを命名した。属名は巨大な猛禽類として表現されるパタゴニアのテウェルチェ族の神話に登場する霊に由来し、種小名は発見者への献名である[3]。
ホロタイプ標本にして唯一既知の標本はほぼ完全な頭蓋骨から構成されており、その上下にやや潰れて眼窩後側の口蓋骨の大部分が失われているものの、眼窩の大部分や頭蓋天井(英語版)や神経頭蓋および方形骨が保存されている。また標本には1個の左足根中足骨(英語版)や、後肢の1個の趾骨の上部、および未同定の骨片が保存されている。これらの骨が共に収集されていてかつ他の骨が混在していないこと、色や質感といった保存状態が全体的に同様であること、また大型フォルスラコス科鳥類のものとして形態学的に調和的であることから、記載者らはこれらの骨が単一の標本のものであると結論した。本標本はその当時において最も完全な大型フォルスラコス科鳥類の頭蓋骨を有した。それ以前に知られていたフォルスラコス科鳥類デヴィケンジア(英語版)やフォルスラコスの頭蓋骨は断片的であり、嘴の先端のみを残す形で破損していたため、ケレンケンが発見されるまで異なる体サイズのフォルスラコス科は分類群間の比較が妨げられていたのである[3]。
進化


標本の大きさや、側方に圧縮されていて強いフック状をなす嘴、よおび凸状の嘴峰に基づき、Bertelli et al. (2007)はケレンケンをフォルスラコス科に分類した。当時はブロントルニス亜科・フォルスラコス亜科・パタゴルニス亜科・メセンブリオルニス亜科・プシロプテルス亜科の5亜科が知られていたが、それらの有効性は分岐分析で確かめられておらず、記載者らは伝統的にフォルスラコス亜科と考えられ板分類群にケレンケンが最も類似するとした。フォルスラコス亜科は後頭部が低く上下に圧縮されていること、後頭部のoccipital tableが広いこと、後眼窩突起(英語版)が鈍いこと、足根中足骨がティタニスのものと類似して下端のsupratrochlear surfaceが平坦であるといった形質を共有する。フォルスラコス亜科の解剖学的情報が不足していたため、さらなる比較は不可能であった[3][5]。
Alvarenga et al. (2011)はフォルスラコス科の系統解析を実施し、ケレンケンとデヴィケンジアが姉妹群であることを突き止めた。またこの解析により5亜科の存在が支持されたが、クラドグラムではブロントルニス亜科・フォルスラコス亜科・パタゴルニス亜科が隔てられなかった[6]。Degrange et al. (2015)はララワビスの記載に際してフォルスラコス科の系統解析を実施し、フォルスラコス亜科が多系統群であることを明らかにした。以下のクラドラムは2015年の解析に従ってケレンケンの系統的位置を示す[7]。
| フォルスラコス科 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
非鳥類型恐竜の絶滅後、新生代の初期において哺乳類が多様化し、鳥類は大型化の傾向を示した。新生代に出現した大型鳥類には、ガストルニス科やドロモルニス科、古顎類、そしてフォルスラコス科がいる[6][8]。フォルスラコス科はノガンモドキ目に属する絶滅したグループであり、当該の目の現生種はノガンモドキ科のアカノガンモドキとクロアシノガンモドキの2種のみとなっている。フォルスラコス科はノガンモドキ目の中でも特異的なグループであるが、化石の不完全性ゆえに、科内における類縁関係は明らかでない[9]。
フォルスラコス科は古第三紀暁新世において当時他の大陸から孤立していた南アメリカ大陸に生息し、第四紀更新世まで生存した。彼らは新第三紀鮮新世の末にはアメリカ大陸間大交差を介して北アメリカ大陸に進出し、また分類が議論されているもののヨーロッパからもフォルスラコス科に分類された化石が報告されている。フォルスラコス科の起源の地は明らかにされておらず、ノガンモドキ科とフォルスラコス科のいずれも南アメリカ大陸で出現したか、あるいは南アメリカ大陸に近接した際に他の地域から進入した可能性がある[5][6]。フォルスラコス科は更新世まで生存していたため、鮮新世に絶滅を迎えた後獣下綱のティラコスミルス科の肉食哺乳類よりも長く命脈を保ったことになり、また鮮新世の間に北アメリカ大陸から進入した有胎盤類の捕食動物と生態的地位を巡って競争していた可能性がある[10]。
ケレンケン自体はフォルスラコス科が絶滅を迎えるよりも遥か以前、まだ肉食性の有胎盤類が南アメリカ大陸へ進出せず後獣類の捕食動物が生息していた時代である、約1500万年前の新第三紀中期中新世(英語版)に生息した[3]。
特徴

フォルスラコス科は大型の飛べない鳥であり、長い後肢と狭い骨盤を持ち、翼がプロポーション的に小型で、巨大な頭骨は前後上下に発達し左右に狭いフック状の嘴を有した[9]。ケレンケンは既知のフォルスラコス科の中で最大であり、フォルスラコスのような従来知られていた最大のフォルスラコス科鳥類よりも10%大型であった。嘴の先端から後頭部に位置する矢状線上の項稜まででホロタイプ標本の頭蓋骨が長さ約71.6センチメートルに達しており、これは既知のいかなる鳥類の中でも最大の頭蓋骨であるとともに、哺乳類のウマのものに匹敵する。頭蓋骨の後端は幅312ミリメートルである。足根中足骨は長さ437ミリメートルである[1][3][8]。現生のノガンモドキが高さ90センチメートルに達する程度であるのに対し、ケレンケンの高さは3メートルにも上った。体重は特に推定されていないものの、100キログラムを超過したと考えられている[8][11]。
頭骨
ケレンケンの発見以前において不完全に知られていた大型フォルスラコス科鳥類の頭蓋骨は、プシロプテルスやパタゴルニスといったより完全な標本が知られている小型の属に基づいて、そのスケールアップ版として復元されていた。例として、1895年に描かれたフォルスラコスの破損した頭蓋骨のスケッチは、パタゴルニスの頭蓋骨に基づいている。これらの復元では上下に非常に高い嘴や、丸みを帯びた上下に高い眼窩、そしてアーチ形天井の神経頭蓋が強調された。しかしケレンケンの発見以降、フォルスラコス科の大型属と小型属との間には頭蓋骨に重大な差異が存在することが示されることとなった[3][4]。ホロタイプ標本の頭蓋骨は非常に巨大で、背側から見ると三角形をなし、後頭部が上下に圧縮されている。上顎の嘴は非常に長く、頭蓋骨の全長の半分を超過しており、メセンブリオルニスやパタゴルニスおよびフォルスラコスよりも長い。骨の鼻孔と前端の距離に基づけば、上顎の嘴と頭蓋骨との比率は0.56である。上下に潰れていることを加味しても、上顎の嘴は高くかつ頑強であるものの、パタゴルニスやアンドリューソルニス(英語版)やアンダルガロルニスほど高く発達してはいないようである[3]。

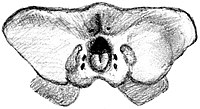
右側(上)と後側(下)から見たホロタイプ標本の頭骨のスケッチ
前上顎骨の前端は鋭利で下側に向いたフックとして顕著に発達する。このような下側の突起は特記の弱い小型のプシロプテルス科と異なり、フォルスラコス、パタゴルニス、アンドリューソルニス、アンダルガロルニスといった中~大型フォルスラコス類に見られる状態と最も類似している。上顎の嘴の下側は1対の発達した稜を形成しており、これが噛縁(英語版)(嘴の左右の縁)から1本の溝によって隔てられ、長軸方向の溝(rostral premaxillar canal)によって前上顎骨の幅広な中央部からも隔てられている。パタゴルニスは口蓋骨の前部に同様の形態を示す。上顎の嘴の大部分は小型の不規則な神経孔が存在する。上顎の嘴の後方2/3は発達した皺が噛縁と平行に走っている[3]。
鼻孔は小型の長方形で、パタゴルニス亜科と同様に上顎の嘴の後背側に位置する(フォルスラコス亜科やブロントルニス亜科において鼻孔の大きさや位置は不明)。鼻孔は上下よりも前後に発達しているが、これは頭蓋骨の変形により誇張されている可能性がある。後縁は鼻骨の上顎突起により形成されている。鼻孔が正中線上で互いに連結していたか、すなわち隔壁が存在しなかったかは識別不能である。前眼窩窓は両側でやや潰れているにも拘わらず四角形であることが明確である。前眼窩窓の前側の縁は鼻孔の後縁と同程度の水準に位置しており、また下縁は左側から見て直線状である。涙骨は頑強であり、前眼窩窓の後縁から伸びており、前頭骨の外側縁や頬骨に対して窪んでいる。前眼窩窓はプロポーション的にパタゴルニスのものよりも小型である[3][4]。

眼窩の形状は僅かに上下の圧縮の影響を受けているものの、上下に低いほぼ長方形の形状であった可能性が高く、また窪んだ上縁と僅かに凸状の下縁を持つ。眼窩の上部は厚い丸みを帯びた縁に縁取られており、その後部が外側から見て下側に突出する。パタゴルニスにおいて同様の構造が涙骨の突起であることが示唆されており、ケレンケンにおいてこれら間の接続が明確でないものの、この構造は涙骨の張り出しであったと推測される。眼窩上部の肥厚は後眼窩突起を形成する前頭骨の一部によって形成されたソケット内に収納されるが、これは他のフォルスラコス科で確認されていない形質状態である。眼窩の下縁は頑強な頬骨の棒状部分によって形成されており、この棒状部分はデヴィケンジアのものよりも上下に発達していてかつ左右に平坦である。頬骨は眼窩の下部中央で高さが厚さの約4倍に発達しており、他のフォルスラコス科を上回る[3]。
前頭骨は背側において平坦である。前頭骨が前上顎骨と接するであろう部分は損傷のため縫合線を特定不可能であるが、前頭骨と鼻骨と頭頂骨との間は完全に癒合している。この癒合により、これらの骨がどのように頭蓋天井の形成に参加しているかの特定が難しいが、鈍く頑強な後眼窩突起はおそらく主に前頭骨により形成されている。左右の前頭骨は下側において筋肉の付着する大型の窪みを持つ。後眼窩突起は頑強な頬骨突起(英語版)により隔てられており、これら2個の突起が狭い1個の側頭窓を閉鎖している。後眼窩突起には強靭な顎の筋肉による筋痕が存在する。頬骨突起の後部には鱗状骨の縁に沿って発達した窪みが存在しており、顎を閉じる筋肉と対応する。より後方のsubtemporal fossaは幅広であり、側方に張り出した鈍い項稜が縁をなす[3]。
上顎骨は長く伸びた口蓋を形成しており、上顎の嘴の長さの大部分に亘ってほぼ平行な縁を持ち、眼窩付近の領域で左右に拡大する。パタゴルニスと同様に、これらの骨はその長さの大部分を走る目立つ長軸方向の窪みによって正中線上で隔てられている。この窪みは口蓋の後側半分に沿って上顎骨の一部に挟まれている。上顎骨後方の側縁は頬骨と縫合されており、パタゴルニスと同様に明瞭である。眼窩後方の頭蓋天井は平坦であり、側頭筋の発達によって筋痕が存在する。デヴィケンジアと同様にoccipital tableは非常に幅広で低いため、後側から見て長方形をなす。後頭顆(英語版)は丸みを帯びており、上面に端を発して顆のほぼ中央部に届く垂直方向の溝を伴う。大後頭孔はほぼ三角形で、上側の頂点が鈍く、顆より僅かに小型である。大後頭孔の直上は稜状に発達した隆起が存在し、大後頭孔の縁から垂直に項稜まで伸びる。顆の下の窩は視認不能であり、窩が目立つパタゴルニスやデヴィケンジアと異なる[3][4]。
後肢

足根中足骨の軸部はある程度細長く、ほぼ中部の断面が長方形であり、フォルスラコスのものと類似する。上面の上側の2/3は窪んでいるが、下側の1/3は平坦である。足根中足骨には軸部の上端にカップ状の2個の臼が存在し、それらはほぼ卵型で深く窪んでいる。外側の臼は内側の臼よりも小型であり、僅かに低い位置に存在する。臼の間隆起はよく発達しており、他のフォルスラコス科と同様である。本属に固有の特徴として、外側の臼の内足底側の隅に丸みを帯びた結節が存在しており、臼間の隆起よりも高さに比べて低い。足根中足骨の軸部の中部は不規則な四角形であり、非常に幅広な長方形をなすブロントルニス亜科のものと異なる。第III中足骨に関節する滑車は第II中足骨や第IV中足骨に関節する滑車よりも遥かに大型であり、より下側に突出している。また、第II滑車と第IV滑車では後者の滑車の方が大型である。第IV滑車は不規則な四角形であり、デヴィケンジアの長方形の滑車と対照的である。足根中足骨の下前側に位置する遠位の血管孔は中心に位置しており、第IIIおよび第IV滑車の直上に位置する[3]。
古生物学
食餌と食性

フォルスラコス科は地上棲の捕食者あるいは腐肉食者と考えられており、また新生代の南アメリカ大陸において優勢であった頂点捕食者と考えられることも多い。当時の南アメリカ大陸に有胎盤類の肉食哺乳類が生息しておらず、代わりにフォルスラコス科はボルヒエナ科に属する大型肉食哺乳類と共存していた。フォルスラコス科の食餌に関する初期の生態学的仮説は主にフック状の嘴を持つ大型の頭骨から類推したものであり、走行性や捕食適応に関する生体力学的研究や詳細な仮説は21世紀初頭以降に行われることになる[9][12]。
Alvarenga and Höfling (2003)はフォルスラコス科の習性について一般的な意見を述べた。フォルスラコス科はその翼の大きさや体重からして飛翔能力を欠いており、またその翼はより大型の個体ほど小型化する。Alvarenga and Höfling (2003)はフォルスラコス科の持つ骨盤・上顎骨上部・胸部の狭いことが背の高い植物や崩れた地勢で小型の獲物の探索・捕獲を可能にしたと指摘した。現生のタカ目と同様に目の上に張り出した涙骨は太陽に対して目を保護して鋭い視力を実現する役割を果たしたと見られ、このことからフォルスラコス科は影の落ちる森でなく太陽に照らされた開けた地で狩りを行ったことが示唆されている[5]。
後肢の機能

Blanco and Jones (2005)は走行速度の推定値が現生の動物においてさえ信頼に足るものでないと懸念を表しつつ、フォルスラコス科の脛足根骨(英語版)の強度を調査して彼らの走行速度を推定した。脛足根骨の強度から推定された走行速度は、未同定の大型フォルスラコス亜科鳥類とパタゴルニスで秒速14メートル、メセンブリオルニスで秒速27メートルで、後者は現生のダチョウ属を上回ってチーターの秒速29メートルに迫っている[13][14]。Blanco and Jones (2005)はこのような高速の推定速度が算出されたことについて、フォルスラコス科の体サイズゆえに蓋然性が低いとし、サイガやトムソンガゼル程度の中型哺乳類の長骨を破壊することにフォルスラコス科自身の骨にも強度が必要であったと推測した。大型でカーブした薄い鉤爪も蹴りを武器として用いたという類推に調和的であり、また自身の骨の硬度を活かして獲物の骨髄を摂食したことも考えられている[13]。
Chiappe and Bertelli (2006)は、フォルスラコス科は大型であればあるほど体がかさばって走行性が低下するという体サイズと俊敏性の電動的な相関関係の考え方について、ケレンケンの発見が一石を投じるものであるとした。ケレンケンの細長い足根足根骨は、より小型で重厚であったブロントルニスよりもケレンケンが俊敏であった可能性を示唆する[4]。この発見に関する2006年のニュース記事にてキアッペは、仮にケレンケンがダチョウ属ほど速くなかったとしても後肢の骨が現生のレア属に類似していることから、従来フォルスラコス科が考えられていたよりも明らかに高速で走行可能であったと述べた。当該記事ではケレンケンが小型哺乳類あるいは爬虫類を追走可能であったとの予想を示唆した[1]。また別の2006年のニュース記事にて、キアッペはケレンケンがイングリッシュ・グレイハウンドと同程度の速度で走行可能であったろうとし、また当時共存していた南アメリカの他の大型捕食動物の走行速度がフォルスラコス科のそれに及ばなかったとした。キアッペの主張によれば、フォルスラコス科はティラノサウルスのような先の時代の肉食性獣脚類との間で大型の頭部・非常に小型の前肢・非常に長い後肢を共通して有しており、同様の肉食適応を遂げていた[15]。
頭部と頸部の機能


Degrange et al. (2010)はコンピュータ断層撮影を用いた有限要素法に基づいて中型フォルスラコス科鳥類アンダルガロルニスの研究を行い、 その咬合力と頭蓋骨の応力分布を推定した。推定された咬合力は嘴の先端部で133ニュートンに達し、またケレンケンのような他の大型フォルスラコス科鳥類と同じく頭蓋内の骨の不動性が大きく失われていることが示された。Degrange et al. (2010)はこの結果について、頭蓋骨の剛性を向上する適応と解釈した。現生のアカノガンモドキやオジロワシと比較してフォルスラコス科の頭蓋骨は横向きの荷重に対して比較的高い応力を示した一方、上下に果汁を加えられた場合や引き戻しのシミュレーションで低い応力を示した。頭蓋骨の側面と正中線が比較的弱いことから、Degrange et al. (2010)はアンダルガロルニスが抵抗する大型の獲物を嘴で鎮圧するような潜在的に危険な行動を取った可能性が低いと結論した。アンダルガロルニスは丸飲みにするなど安全に殺害して消費可能な小型の獲物を捕食したか、あるいは嘴を用いてヒット&アウェイ戦法で大型の獲物を標的にしたことが提唱された。抵抗する獲物を足で抑え込むことも可能であった[12]。
Tambussi et al. (2012)はアンダルガロルニスの頸椎の形態に基づいてその頸部の柔軟性を解析し、頸部を3つのセクションに区分可能であることを発見した。頸椎を手動で操作することにより、Tambussi et al. (2012)はアンダルガロルニスの頸部の筋肉と骨格が大型の頭部を運ぶことに適応し、また下向きの打撃後に最大延長から持ち上げることの補助に適応していると結論した。さらに、この結論は大型の頭部を持つ他の大型フォルスラコス科鳥類にも当てはまると類推された[16]。Degrange (2020)によるフォルスラコス科のお頭蓋骨形態の研究では、ノガンモドキ型の祖先から派生した2つのモルフォタイプが存在することが明らかにされた。1つは原始形質である"Psilopterine Skull Type"、もう1つはケレンケンや他の大型属を含む、頭蓋骨がより頑強に特殊化した"Terror Bird Skull Type"である。このような差異があるものの2種類は獲物を同様の方法で取り扱っていたが、より強固な頭蓋骨とより強大な咬合力を持つ"Terror Bird Skull Type"はより大型の獲物を対象とする適応を遂げていたと考えられている[9]。
古環境

ケレンケンはパタゴニア地域のコマロの南東部に分布するCollón Curá 層に属する火山砕屑岩の露頭から産出しており、当該地域は白色がかった凝灰岩に被覆されている。当該地域の層序は発見当時において予察的に研究されていた程度であり、堆積物の年代も十分に決定されていなかった。Collón Curá層の異なる地域から得られた放射年代測定の結果と南アメリカの陸生化石動物の年代との対比から、ケレンケンの産出層準は約1500万年前にあたる中期中新世と推定された[3][8][17]。本層は互いに不連続な複数の構造盆地を特徴とする崩壊した前縁盆地(英語版)で堆積した。本層は火山砕屑岩や石灰岩および砂岩を主体としており、堆積環境は流水や湖沼といった淡水圏の大陸環境であった[18]。
Collón Curá層の堆積期間において、当該地域は植生による被覆が減少したより開けた環境が優勢であり、半乾燥気候から温帯気候の低木林地(英語版)や森林地帯に類似した環境であったとされる。開けた環境は走行に適した動物や大型動物の出現に繋がり、樹上性の動物が生息する発達した森林が存在した前期中新世(英語版)後期の序盤の状態と対照的であった。森林は山系の渓谷に限定され、樹上性の種も僅かであったとされる。この変化は南アメリカの地質時代であるフリアシアン(英語版)期前期の間に発生して進行したと見られる[19][20]。乾燥した景観への移行は、中期中新世に発生した全球的寒冷化に対応する気候変動と同時に発生したと見られる[20]。
Collón Curá層は様々な哺乳類群集の化石が算出しており、異節類(Megathericulus、Prepotherium、Paraeucinepeltusなど)、南蹄目のProtypotherium、雷獣目のアストラポテリウム(英語版)、砕歯目のパタゴスミルス(英語版)やクラドシクティス(英語版)、有袋類のAbderites、霊長目のProteropithecia、齧歯目(Maruchito、Protacaremys、Neoreomys、Prolagostomus)といった24分類群以上の哺乳類が報告されている[17][18][21][22]。哺乳類以外にも鳥類・爬虫類・両生類・魚類の化石が産出している[19]。
出典
- ^ a b c Wilford, John Noble (31 October 2006). “Fossil found of a Big Bird Kermit wouldn't like”. The New York Times. 26 May 2022閲覧。
- ^ Hotz, Robert Lee (26 October 2006). “Teen finds fossil skull of biggest bird known”. Pittsburgh Post-Gazette: p. 13. http://news.google.com/newspapers?nid=1129&dat=20061026&id=44dIAAAAIBAJ&sjid=YHIDAAAAIBAJ&pg=6604,1821231 26 May 2022閲覧。
- ^ a b c d e f g h i j k l m n o p q Bertelli, Sara; Chiappe, Luis M.; Tambussi, Claudia (2007). “A new phorusrhacid (Aves: Cariamae) from the middle Miocene of Patagonia, Argentina”. Journal of Vertebrate Paleontology 27 (2): 409–419. doi:10.1671/0272-4634(2007)27[409:ANPACF]2.0.CO;2. https://www.researchgate.net/publication/249023842.
- ^ a b c d e Chiappe, Luis M.; Bertelli, Sara (2006). “Skull morphology of giant terror birds”. Nature 443 (7114): 929. doi:10.1038/443929a. PMID 17066027. https://www.researchgate.net/publication/6730231.
- ^ a b c Alvarenga, Herculano M.F.; Höfling, Elizabeth (2003). “Systematic revision of the Phorusrhacidae (Aves: Ralliformes)”. Papéis Avulsos de Zoologia 43 (4). doi:10.1590/S0031-10492003000400001.
- ^ a b c Alvarenga, Herculano; Chiappe, Luis; Bertelli, Sara (2011). “Phorusrhacids: The Terror Birds”. In Dyke, Gareth; Kaiser, Gary. Living Dinosaurs. pp. 187–208. doi:10.1002/9781119990475.ch7. ISBN 978-1-119-99047-5
- ^ Degrange, Federico J.; Tambussi, Claudia P.; Taglioretti, Matías L.; Dondas, Alejandro; Scaglia, Fernando (2015). “A new Mesembriornithinae (Aves, Phorusrhacidae) provides new insights into the phylogeny and sensory capabilities of terror birds”. Journal of Vertebrate Paleontology 35 (2): e912656. Bibcode: 2015JVPal..35E2656D. doi:10.1080/02724634.2014.912656. hdl:11336/38650. https://www.researchgate.net/publication/275272037.
- ^ a b c d Ksepka, Daniel T. (2014). “Flights of fancy in avian evolution”. American Scientist 102 (1): 39. doi:10.1511/2014.106.36. ISSN 0003-0996. JSTOR 43707746.
- ^ a b c d Degrange, Federico J. (2020). “A revision of skull morphology in Phorusrhacidae (Aves, Cariamiformes)”. Journal of Vertebrate Paleontology 40 (6): e1848855. Bibcode: 2020JVPal..40E8855D. doi:10.1080/02724634.2020.1848855. https://www.researchgate.net/publication/348614159.
- ^ Tambussi, Claudia; Ubilla, Martín; Perea, Daniel (1999). “The youngest large carnassial bird (Phorusrhacidae, Phorusrhacinae) from South America (Pliocene-Early Pleistocene of Uruguay)”. Journal of Vertebrate Paleontology 19 (2): 404–406. Bibcode: 1999JVPal..19..404T. doi:10.1080/02724634.1999.10011154. ISSN 0272-4634. JSTOR 4524003. https://www.jstor.org/stable/4524003.
- ^ Tambussi, Claudia P.; Degrange, Federico J. (2013). “Neogene Birds of South America”. South American and Antarctic Continental Cenozoic Birds. Springer Briefs in Earth System Sciences. Springer. pp. 59–86. doi:10.1007/978-94-007-5467-6_7. ISBN 978-94-007-5466-9
- ^ a b Degrange, Federico J.; Tambussi, Claudia P.; Moreno, Karen; Witmer, Lawrence M.; Wroe, Stephen (2010). “Mechanical analysis of feeding behavior in the extinct "terror bird" Andalgalornis steulleti (Gruiformes: Phorusrhacidae)”. PLOS ONE 5 (8): e11856. Bibcode: 2010PLoSO...511856D. doi:10.1371/journal.pone.0011856. PMC 2923598. PMID 20805872. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2923598/.
- ^ a b Blanco, Rudemar Ernesto; Jones, Washington W. (2005). “Terror birds on the run: a mechanical model to estimate its maximum running speed”. Proceedings of the Royal Society B: Biological Sciences 272 (1574): 1769–1773. doi:10.1098/rspb.2005.3133. PMC 1559870. PMID 16096087. https://www.researchgate.net/publication/7664536.
- ^ Hudson, Penny E.; Corr, Sandra A.; Wilson, Alan M. (2012). “High speed galloping in the cheetah (Acinonyx jubatus) and the racing greyhound (Canis familiaris): spatio-temporal and kinetic characteristics”. Journal of Experimental Biology 215 (14): 2425–2434. doi:10.1242/jeb.066720. PMID 22723482.
- ^ Joyce, Christopher (25 October 2006). “Huge "terror bird" fossil discovered in Patagonia”. NPR.org. 26 May 2022閲覧。
- ^ Tambussi, Claudia P.; de Mendoza, Ricardo; Degrange, Federico J.; Picasso, Mariana B. (2012). “Flexibility along the neck of the Neogene terror bird Andalgalornis steulleti (Aves Phorusrhacidae)”. PLOS ONE 7 (5): e37701. Bibcode: 2012PLoSO...737701T. doi:10.1371/journal.pone.0037701. PMC 3360764. PMID 22662194. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3360764/.
- ^ a b Kramarz, Alejandro; Garrido, Alberto; Bond, Mariano (2019). “Astrapotherium from the Middle Miocene Collón Cura Formation and the Decline of Astrapotheres in Southern South America”. Ameghiniana 56 (4): 290. doi:10.5710/AMGH.15.07.2019.3258.
- ^ a b Brandoni, Diego; Ruiz, Laureano González; Bucher, Joaquín (2020). “Evolutive implications of Megathericulus patagonicus (Xenarthra, Megatheriinae) from the Miocene of Patagonia Argentina”. Journal of Mammalian Evolution 27 (3): 445–460. doi:10.1007/s10914-019-09469-6.
- ^ a b Tonni, Eduardo P.; Carlini, Alfredo A. (2008). “Neogene vertebrates from Argentine Patagonia: their relationship with the most significant climatic changes”. In Rabassa, Jorge. The Late Cenozoic of Patagonia and Tierra del Fuego. Elsevier Science. pp. 269–278. ISBN 978-0-08-055889-9
- ^ a b Genise, Jorge F.; Bellosi, Eduardo S.; Cantil, Liliana F.; González, Mirta G.; Puerta, Pablo (2022). “Middle Miocene climate transition as reflected by changes in ichnofacies and palaeosols from Patagonia, Argentina”. Palaeogeography, Palaeoclimatology, Palaeoecology 594: 110932. Bibcode: 2022PPP...59410932G. doi:10.1016/j.palaeo.2022.110932.
- ^ Kay, Richard Frederick; Johnson, Derek; Meldrum, Don Jeffrey (1998). “A new pitheciin primate from the middle Miocene of Argentina.”. American Journal of Primatology 45 (4): 317–336. doi:10.1002/(SICI)1098-2345(1998)45:4<317::AID-AJP1>3.0.CO;2-Z. PMID 9702279. https://www.researchgate.net/publication/13583562.
- ^ Echarri, Sebastian; Ulloa-Guaiquin, Karen S.; Aguirrezabala, Guillermo; Forasiepi, Analia M. (2021). “Cladosictis patagonica (Metatheria, Sparassodonta) from the Collón Cura Formation (Middle Miocene), Río Negro, Argentina”. Ameghiniana 58 (6). doi:10.5710/AMGH.06.08.2021.3439. hdl:11336/171288.







